


自然群落里为什么存在形态各异的物种?这些物种又是怎样共存的?生态学家致力于解答这些疑问, 并由此提出了众多解释物种共存的理论和假说(Wright, 2002)。比如, 种库理论、更新生态位理论、资源比率/异质性假说、竞争共存理论、生态漂变假说等(Grubb, 1977; Tilman, 1982; Taylor et al., 1990; Hubbell, 2001; 侯继华和马克平, 2002)。目前研究者主要围绕两个方面来开展物种共存机制的相关研究: (1)检验这些理论或假说所对应的物种多样性维持机制是否存在; (2)这些机制在自然群落里表现的强度如何(Wright, 2002)。如果在自然群落没有检验到某一维持机制的存在, 或者该种机制对群落组成的影响较小, 那么就可以认为这一理论或假说不能或者不足以解释物种共存; 如果检验到某一维持机制影响群落结构, 并在群落里广泛存在, 那么可以认为它是维持物种多样性的主要机制。植物自身的特性和环境条件是检验这些理论或假说的基础。
其中两个从不同角度解释物种共存的假说得到了有力支持。一个是不同物种通过占有不同空间和资源实现共存的生态位分化理论; 另一个是负密度制约假说, 即由于资源竞争、有害生物侵害(比如病原微生物、食草动物等)和化感作用等导致同种个体之间相互损害, 从而为其他物种的生存提供空间和资源, 促进物种共存。学者们对这两个主要的理论提出了不同的观点(Condit et al., 1992; He et al., 1997; Hubbel, 1997; Hyatt et al., 2003)。比如He等(1997)认为生态位理论和负密度制约虽然都能影响群落的物种组成, 但两者对不同的物种或物种的不同生活史阶段等所起作用不同。而Hubbell(2001)认为两者都不足以解释群落里多种物种的共存, 进而提出中性理论。他假定不同物种、不同个体在生态功能上完全等同, 具有相同的出生率、死亡率和扩散率, 物种灭绝、物种迁入和新物种形成的动态平衡决定群落中的物种数。
本文将主要介绍物种共存的负密度制约假说, 有关这一假说的概念发展、研究的主要方向及其进展等, 希望有助于大家深入理解和探讨这方面的科学问题。
1 负密度制约假说的概念发展
20世纪30年代, Ridley(1930)指出自然群落中大量同种个体生长在一起, 易受到有害生物(比如害虫、菌类等)的侵害。50年代, 生态学家们发现热带森林里每个物种的个体分布, 与随机过程相比表现得更为规则(Black et al., 1950; Pires et al., 1953)。70年代, Janzen (1970)和Connell (1971)分别撰文, 系统地描述了负密度制约能够解释热带森林维持较高物种多样性的机理, 形成了Janzen-Connell假说。
Janzen-Connell假说认为种子扩散以母株为中心, 邻近母株的种子和幼苗存在较高的死亡率, 这有助于维持热带森林物种多样性。该假说主要包括两个过程: (1)大部分植物的种子落在母株周围,种子数量常随着远离母株的距离而下降, 而且随着扩散期间种子产量的波动而变化; (2)母株和其种子、幼苗是许多宿主专一的植物病原菌和捕食者的食物来源, 这些有害生物导致更多邻近母株的种子和幼苗死亡, 它们对更新后代的负效应随着远离母株而下降, 而幼苗更新存活的机率随着远离母株而增加(图1)。这两个过程的相互作用产生了一个距母株一定距离的种群增补曲线(population recruitment curve), 在曲线范围内, 种子得以萌发, 部分幼苗能够长成大树, 总体更新个体数量在远离母株一定的距离处达到最多。Janzen(1970)认为大部分森林具有多物种共存、多数物种的密度低、成年树分布较规则的特点, 这是上述两个过程共同作用的结果。
图1
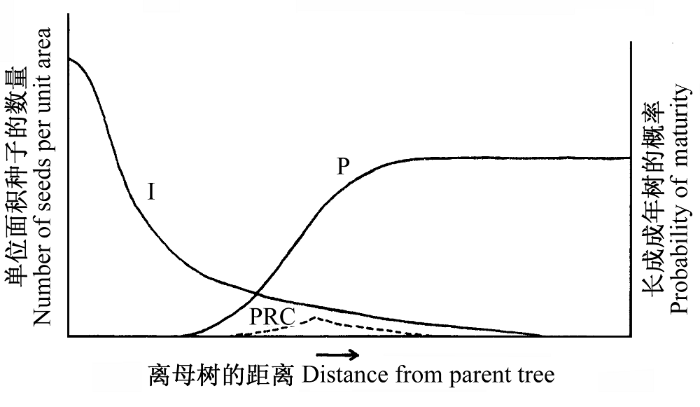
图1
Janzen-Connell假说中种子扩散和距离制约种子、幼苗死亡的过程(Janzen, 1970)。PRC: 种群增补曲线; I:单位面积种子数量; P: 种子或幼苗幸免于捕食的概率。
Fig. 1
The processes of Janzen-Connell hypothesis on seed dispersal and distance dependence on seed or seedling mortality (Janzen, 1970). PRC, Population recruitment curve;I, The number of seeds per unit area; P, The probability that a dispersed seed or seedling will be missed by the host-specific seed and seedling predators.
基于这两个过程, Janzen(1970)提出了关于热带森林物种多样性维持的一般性假说, 包括5点: (1)种子萌发长成小树的成活率随着离母株距离的增加而提高; (2)种子扩散前在母株上的死亡率和周围成年树之间的距离呈负相关; (3)种子扩散前的死亡率和繁殖母株的密度呈正相关; (4)任何一株小树的成活率和它周围同种小树的数量呈反比; (5)符合 (1)和(4)假说的种群, 其成年树的分布格局为均匀 分布。
按照Janzen-Connell假说, 由于有害生物从母株传播到附近的增补后代(recruitment offspring)上, 聚集的宿主有利于专一性草食动物和病原体的繁殖和传播, 使得同种邻体表现为较低的生长率和存活率。遭受侵害的物种, 其同种个体的间距变大, 将会腾出空间和资源给与它天敌不同但有相似 资源需求的其他种, 从而提高热带森林的物种多 样性。
早期, 因为解释物种共存的假说较少, 为了更好地理解为什么森林里有多个物种共存, 许多科学家致力于探讨Janzen-Connell假说的有效性(Hubbell, 1979; Howe & Smallwood, 1982; Condit et al., 1992)。后来, 研究者把有关Janzen-Connell假说的各种密度和距离制约效应统称为同种之间的负密度制约, 简称密度制约。此外, 负密度制约是描述同种个体之间相互作用的宽泛的概念, 包含文献上较常提到的负频率依赖(negative frequency dependence)(Wills et al., 1997)。负频率依赖常以物种同种个体数量发生的变化为参数来研究负密度制约效应是否发生, 而负密度制约可以用多种参数形式来研究, 比如生物量的变化等, 也包括同种个体的数量变化(Wills et al., 1997)。
负密度制约假说与中性理论存在较大差异。中性理论假设群落内各物种在生态功能上等同, 同种邻体之间的竞争和异种邻体之间的竞争相同 (Hubbell, 1997; Hubbell, 2001)。而负密度制约模型则假设物种之间是不同的, 由于对相同资源的竞争, 或者由于病原菌、食草动物等的侵害, 个体在同种密度高的生境中存活率较低; 相反, 如果个体出现在异种密度高的生境, 由于同种个体之间对相同资源的竞争减弱, 受病原菌感染和食草动物的捕食较弱, 个体的成活率较高。
2 负密度制约发生的机理
植物是固着的自养型生物, 必须在原地繁衍生息, 既要承受来自其他生物的侵害, 又要遭受邻体对水分、矿质营养和光照的竞争(Wright, 2002)。同种个体之间主要有两大类因素能够导致负密度制约效应的发生。第一, 有害生物对植物体的侵害。主要指某一营养级的生物(食草动物、病原微生物等)对另一营养级的生物(植物)的取食, 种群密度高的个体更易受到损害(Janzen, 1970; Connell, 1971)。 第二, 资源竞争。指的是同一营养级之间竞争资源, 导致个体之间相互损害。同种个体具有同样的生长习性和资源需求, 由于资源有限, 同种邻体之间发生资源(光照、营养、空间等)竞争, 这样就可导致同种个体生长量下降、死亡率升高 (Comita & Hubbell, 2009)。
2.1 有害生物侵害
Janzen (1970)和Connell (1971) 假设病原菌、食草动物等能够降低邻近同种成年个体的更新率和生长率, 从而给予其他物种生存的空间和资源, 促进物种共存。许多对于单一物种的研究已经发现有害生物能够导致负密度制约的发生(Augspurger, 1984; Gilbert et al., 1994; Gilbert & DeSteven, 1996; Packer & Clay, 2000; Bell et al., 2006)。Hammond和Brown(1998)综合分析了46个有关Janzen-Connell假说的研究案例, 比较了不同类型有害生物对成年个体周围不同距离上种子或幼苗的影响。他们发现大部分昆虫类食草动物能够降低紧邻成年个体的更新成功率, 但是脊椎动物研究案例中极少出现这种现象。在中国海南岛尖峰岭热带低地雨林和季雨林, Li等(2009)发现聚集在软荚红豆(Ormosia semicastrata)母体周围的专一宿主病原菌可以调节其幼苗阶段的群落结构。在母体周围的土样处理中, 他们发现土壤灭菌与否显著影响软荚红豆种子萌发率和幼苗死亡率: 在灭菌土壤里的种子萌发率较高, 幼苗死亡率较低。然而, 在远离母体的土壤里, 幼苗死亡率和成活率不受土壤灭菌处理与否的影响。也就是说, 目前的研究基本认同昆虫和少量病原微生物对植物体的影响符合Janzen-Connell假说。然而, 还不清楚在森林群落范围内昆虫和病原体是否能降低大部分物种的更新成功率(Wright, 2002)。
虽然大量证据说明有害生物对森林植物的捕食压力能促进热带森林的物种共存(Leigh et al., 2004), 但是有害生物与负密度制约发生的关联性研究仍然较少。Carson等(2008)综合分析了50项符合密度或距离制约预测的研究, 发现其中50%的研究提供了发生负密度制约机理的证据, 包括8项脊椎动物、10项无脊椎动物和7项病原菌的研究, 然而另外50%的研究没有提供有害生物与负密度制约发生的关联性证据。此外, 虽然Janzen和Connell都认为宿主专一性是负密度制约发生的重要因素, 但是只有较少的研究探讨过这一问题(Carson et al., 2008)。另外, 在哥斯达黎加La Selva生物学研究站, 研究者分析了热带森林的土壤病原体与幼苗各种生长参数的关系, 发现土壤病原体不是通过负密度制约效应调节群落结构, 而是通过促进生态位分化来达到多物种的共存(McCarthy-Neumann & Kobe, 2008)。而且, 如果说有害生物是负密度制约的因子, 那么有害生物捕食专一化程度仍需量化。
2.2 资源竞争
资源匮乏导致同种个体竞争激烈, 较多的文献报道了关于光资源竞争的负密度制约研究。研究者一般把研究对象界定为喜阳或耐荫植物, 比较分析耐荫性与生长率、成活率等各项指标变化的关联性(Hubbell et al., 2001; Comita & Hubbell, 2009)。Hubbell等(2001)在巴拿马BCI(Barro Colorado Island)样地发现: 喜阳树种的个体增长量与成活率呈正相关, 当其高度超过周围的邻体时, 成活率提高159%; 然而耐荫树种的成活率与个体的增长量只有轻微的正相关, 当耐荫树种高度超过它周围的邻体时, 成活率仅提高4%; 喜阳树种和耐荫树种都与同种邻体密度呈负相关, 但是喜阳树种对同种邻体的敏感程度高于耐荫树种; 并且在同样的光资源下, 喜阳树种的个体死亡率高于耐荫树种。
资源短缺常成为植物生存的限制因素, 同种邻体之间的资源竞争有可能是负密度制约发生的主要因素。然而各种资源类型较难量化, 负密度制约与资源竞争的关联性研究仍相对较少。特别是水分、土壤养分等资源类型结合负密度制约的研究较少开展, 而不同区域水分限制、土壤贫瘠常成为植物生长的主要限制因子, 因此这方面的研究将有助于深入理解负密度制约发生的机理。
此外, 众多研究假设种内竞争是负密度制约发生的基础(Silva Matos et al., 1999; Stoll & Newberry, 2005), 然而负密度制约的发生也是多因子交互作用的结果。比如食草动物啃食乔木幼苗强度大, 会导致幼苗的耐荫性降低, 这样在幼苗没有达到林冠层高度时, 由于光资源的短缺, 大量幼苗将会死亡(Carson et al., 2008)。
3 负密度制约研究的主要方向
一般研究采用种群个体的存活率、生长率和更新率参数检验负密度制约(Wright, 2002)。大量研究发现, 幼苗的密度高或者邻近母体, 死亡率就会高 (如Augspurger, 1983a, b, 1984; Clark & Clark, 1984; Schupp, 1988, 1992; Howe, 1990; Gilbert et al., 2001)。负密度制约研究主要聚焦在4个方面: (1)种群密度大小与负密度制约发生的关系。通过比较样方内目标物种(focal species)的密度与个体死亡率等参数的关系, 检验同种个体密度的大小是否影响种群结构的变化(Wills et al., 1997)。 (2)更新后代离母株距离的远近与负密度制约发生的关系。有害生物从母体传播到邻近的种子、幼苗和幼树上, 产生自上而下的损害, 通过分析种子的萌发率、幼苗的更新率、生长率等与母株距离的关联性来检验距离制约效应的存在(Condit et al., 1992)。(3)群落补偿趋势(community compensatory trend, CCT)。负密度制约效应发生的强度与种群大小呈正相关的关系, 这种趋势有利于稀有物种在群落尺度上的存活、更新和生长, 促进群落水平的物种共存(Connell et al., 1984)。(4)研究不同物种个体之间的关系, 属于负密度制约研究的新扩展。比如异种邻体可能阻碍有害生物的传播, 据此建立了促进物种共存的物种异群保护模型(species herd protection)(Peters, 2003)。 Webb等(2006)引入物种之间的谱系关系(phylogeny), 提出了谱系多样性模型(phylodiveristy dependence)。
3.1 种群密度大小与负密度制约
早期的研究只发现少数丰富种(abundant species)表现出负密度制约现象。因此, 一般认为负密度制约不是群落内维持生物多样性的普遍性机制(Hubbell, 1979; Hubbell et al., 1990)。Hubbell和Foster(1986b)采用模型J = a0+a1A+a2A2(a0, a1和a2是常数, J和A分别代表小树和成年树的密度), 拟合50个1 ha样地的48个丰富种的小树和成年树的密度, 发现只有最丰富的物种表现出强烈负密度制约效应(a1<0)。在巴拿马BCI样地, 研究者只发现最丰富物种鹧鸪花(Trichilia tuberculata)的种群密度受到负密度制约效应的调节(Hubbell et al., 1990)。还有一些物种, 比如阿沙依椰子(Euterpe edulis)、巴西棕榈(Astrocaryum mexicanu)和钝叶蚁栖树(Cecropia obtusifolia), 其植株生长速度不受周围种群密度大小的影响(Alvarez-Buylla, 1994; Alvarez-Buylla et al., 1996; Silva Matos et al., 1999)。Wills和Condit (1999)在对巴拿马和马来西亚热带森林样地的研究中甚至发现, 胸径大于1 cm的树木死亡呈现随机性。
早期研究者较少认为负密度制约是森林群落物种共存的普适性调节机制, 主要原因有三方面:
第一, 早期检验负密度制约效应一般采用样方分析的方法, 即把大样方分为多个小样方, 比较每个样方中相对密度和死亡率的差别, 但是样方边界效应会导致树木真实的邻体信息丢失, 从而低估负密度制约的作用(Hubbell et al., 1990)。正如Peters(2003)所述, 早期研究普遍采用边长为5-20 m的方形样方, 邻近样方边界(离边界距离小于2 m)的个体占总株数的36-96%。因此, 采用这种方法将会丢失大部分树木的邻体信息, 难以检测到负密度制约的发生。
第二, 空间资源的异质性将会掩盖潜在的负密度制约效应发生的迹象。在适合植物生长的生境, 负密度制约所导致的个体死亡或者生长量减少, 被由于生境适宜导致的成活率提高和个体生长量的增加所抵消。在适于种群生长的生境不仅难以检验出负密度制约效应, 反而会得到正密度制约效应的假象。
第三, 难以排除的自然干扰。比如暴风雨、火灾等大的自然灾害导致的植物个体死亡, 以及植物自身种子产量、生长量和死亡率的年际变化, 都会掩盖负密度制约的作用(Condit, 1995; Wright et al., 1999; Wright, 2002)。
之后, 研究者改进了方法, 检测到更多物种受到负密度制约效应的影响。Wills等(1997)发现在BCI样地研究的84个物种中有67个表现出显著的负密度制约更新效应, 这一效应已成为维持物种多样性的主要因素, 远比种间效应更普遍; 同时, 负密度制约显著影响了54个物种的内禀增长率。在巴拿马BCI和马来西亚Pasoh热带森林, Peters(2003)改进分析方法, 消除了小样方边界的影响, 分析了同种邻体聚集对建成树(established tree, 胸径≥ 1cm)死亡率的影响, 发现每个样地内都有超过80%的物种的个体死亡率受到负密度制约效应调节。此外, Zhu等(2009)排除了生境异质性的干扰, 对古田山24 ha森林样地的研究表明, 83%的被检验物种的种群结构受到负密度制约的调节, 进一步证明了负密度制约调节森林群落种群结构的重要性。在BCI样地, Harms等(2000)做了一项负密度制约对更新个体(从种子到幼苗生长过渡阶段)影响的分析。他们发现4年内, 到达200个种子收集器的386,027个种子与邻近样方13,068株幼苗相比, 幼苗种类显著多于种子种类, 说明在从种子过渡到幼苗的过程中, 负密度制约效应普遍提高了物种多样性。
3.2 负密度制约强度与母株距离的关系
密度效应和距离效应对种群结构的影响有时是很难分开的。已有研究表明目标个体同时受到种群密度和同种个体之间距离的影响(Hubbell, 1980; Connell et al., 1984; Hubbell et al., 1990; Gilbert et al., 1994)。距离效应的研究侧重探讨是否紧邻母株的更新后代存活率低, 而种群密度则制约着同种个体的变化, 如同种邻体密度的大小制约个体的生长、存活或死亡, 从而影响种群大小。捕食者(如昆虫、病原体等)易于发现和捕食聚集在母体周围的种子、幼苗和幼树。因此, Janzen-Connell假说预测, 种子发展到建成树的成活率随着与母株距离的增加而提高, 在远离母株一定的距离处, 总体更新个体数量最多(Janzen, 1970)。
距离制约预测是Janzen-Connell假说最基本的部分, 得到研究者广泛的验证(Hubbell & Foster, 1986b; Wright, 2002)。其中Howe和Smallwood (1982)的研究表明: 后代远离母株能够提高更新成功率(逃逸假说); 更新后代通过母株扩散到种群新定居的林窗, 由于林窗中成年树密度低、捕食者类群发生变化, 幼树被捕食的总体机率降低(建群假说)。他们不仅在热带地区而且在温带地区也发现了距离制约效应存在的证据, 扩大了距离制约预测得以验证的区域范围。
距离制约效应对植物不同生长阶段影响的程度不同: 一般幼苗受到的损害最大, 随着个体的生长, 影响逐渐降低。Connell (1971)通过观测研究发现, 种子死亡与母株的距离关系并不紧密, 而幼苗比种子表现出更强的距离制约效应。Hyatt等(2003)综合分析了距离制约的研究案例, 也发现幼苗比种子更能支持距离制约预测。幼苗个体较大, 捕食者较易通过母株发现它们, 而且由于幼苗自身积累的资源有限, 一经捕食者破坏, 便难以正常生长甚至死亡。因此, 幼苗阶段遭受有害生物损害的程度最大, 随着植物个体高度增加, 捕食效应逐渐降低(Connell, 1971; Connell et al., 1984; Condit et al., 2000; Wright, 2002)。由于邻近母株更新个体死亡率高, 故随着时间的推移, 同种树木个体的间距增大, 成年树向更加规则的分布格局发展。
距离制约预测在很多单一物种分析中得以验证, 但其普遍调节森林群落种群结构的证据仍很少。Hubbell(1980)通过检验距离制约效应对于维持物种多样性的潜在作用, 发现树木个体呈现规则分布格局仅能解释少部分物种的共存, 距离制约效应解释森林群落物种多样性维持机制的可能性比较低。Condit等(1992)检验了巴拿马热带雨林BCI样地的80个物种, 发现受到邻近母株距离制约更新效应影响的物种较少, 并且距离制约效应仅发生在离母株小于5 m的范围内。Hyatt等(2003)综合分析了距离制约效应的研究案例, 发现远离母株的生境并不能显著提高更新后代的成活率, 林窗或者不同于母株生长的生境一般也不能提高更新率; 而且, 在温带地区, 随着与母株距离增加, 更新后代的成活率反而稍微降低。由于没有支持距离制约预测的普遍性证据, Hyatt等(2003)甚至认为, 没有必要进一步探讨距离制约效应是否是维持物种多样性的重要机制。同时, Hyatt等(2003)也发现许多物种邻近母株的更新后代有生长量降低、死亡率提高的现象, 承认距离制约效应对一些物种种群结构的调节是重要的, 但是他们认为距离制约效应不是影响种群所有生活史阶段以及所有生活型的调节机制, 更不是普遍存在的群落生物多样性维持机制。但是Wright (2002)和Petermann等(2008)认为包括BCI样地的大多数研究, 均低估了距离制约效应对群落的普遍影响。
距离制约效应在群落中的普遍性被低估的原因主要有以下几方面: 第一, 邻近母株更新后代受到的负效应影响具有滞后性。如果经过更长时间的积累, 距离制约对植物生长和存活的负效应极有可能加强, 并最终引起母体周围更新个体的减少。然而, 大多数的研究时间较短, 很难发现距离制约发生的迹象。第二, 种子扩散限制。种子通常落在紧邻母株的周围, 到达距离母株较远位置的机率低, 因而很可能使邻近母株的更新个体数最多。如果只是分析邻近或远离母株处存活的建成树的数量, 就会很大程度上低估距离制约效应的作用。第三, 适宜的生境抵消了距离制约效应的影响(Wright, 2002)。母株所在的位置一般是适宜种群生长的生境(Hamill & Wright, 1986), 适宜的生境能够提高更新个体的成活率, 如果成活率提高量大于由于距离制约效应导致的个体死亡量, 就会出现邻近母株后代更新率反而提高的现象。而远离母株的生境本身存在不适合种群生长的因素, 不适宜的生境能够降低更新个体的成活率。如果生境不适宜带来的影响高于远离母株的成活率提高的好处, 就可能出现后代远离母株更新率反而降低的现象。因此, 距离制约效应调节种群结构的效应应该比现在已经意识到的更强, 可能是物种多样性维持的一种重要机制。
3.3 群落补偿趋势
群落补偿趋势在群落尺度上描述了负密度制约效应和种群大小之间的密切关系, 即种群数量越大受负密度制约的影响越大, 反之越小。这一概念由Connell等(1984)提出, 主要强调群落补偿趋势能够阻止丰富种以牺牲稀有种为代价扩大生存环境范围, 保护稀有物种免于局部灭绝, 从而促进物种共存。Welden等(1991)在对BCI样地, Webb和Peart (1999)在对婆罗洲150 ha森林幼苗存活更新过程的研究中, 检测到了群落补偿趋势, 而且证明负密度制约是群落补偿趋势发生的机制。Queenborough等(2007)在25 ha的热带森林的研究中发现, 越丰富的物种幼苗死亡率越高, 并且物种之间的微生境和生活史策略的差异不是导致群落补偿趋势发生的原因。然而, He等(1997)在马来西亚Pasoh森林样地的研究表明, 随着种群大小的增加, 小树的成活率提高, 与群落补偿趋势相反, 说明群落补偿趋势也不能解释所有物种的种群动态。
在大部分热带森林, 林窗有利于先锋树种的更新、生长和存活(Hubbell & Foster, 1986b; Denslow, 1987; Brown & Whitmore, 1992)。但在BCI样地经历了严重的El Niño干旱后, 树木死亡率增加, 倒木形成的林窗也增加, Welden等(1991)反而发现了促进稀有种或偶见种更新的群落补偿趋势, 提高了稀有先锋树种占有生境的机会。然而, 另有研究表明稀有物种在种群建立方面表现出相应的弱势(Hubbell & Foster, 1986a)。比如Zhu等(2009)在古田山的研究中发现, 由于物种自身生物学特性的限制, 个体数少的物种比个体数多的物种表现出更强的同种聚集度(如红淡比Cleyera japonica, n = 479), 负密度制约的强度也较大; 而个体数多的物种同种个体聚集度低, 在研究样地中分布均匀, 比如石栎(Lithoc- arpus glabra, n = 1,309)和毛花连蕊茶(Camellia fraterna, n = 4,108), 二者都是丰富种, 分析中没有检验到它们受负密度制约的影响; 总体上个体数多聚集度又高的物种受负密度制约的影响也大(如马尾松, Pinus massoniana)。
Zimmerman等(2008)总结分析了物种共存理论, 认为负密度制约和中性理论最关键的差别在于二者对稀有种在群落里的发展趋势作出了不同的预测。负密度制约假说认为稀有种有生存的优势, 随着时间的推移, 稀有种可能逐渐成为常见种; 而中性理论预测稀有种与丰富种相比在群落里有逐渐消亡的危险(Volkov et al., 2005)。Wills等(2006)分析了7个森林动态监测样地的数据, 发现稀有种的更新优势提高了群落的物种多样性。在局部区域, 现存的群落比更新初级阶段或者死亡个体组成的群落物种多样性高; 树龄较大的群落比树龄较小的群落物种多样性高; 稀有物种比常见种的成活率高, 稀有种的生长优势导致生态系统中存在丰富的物种。他们的工作清楚地证明了稀有种在热带森林中的生存优势。但是稀有种所表现的更新率高、生长快、存活率高等各种优势有可能与微环境紧密相关, 未来群落补偿趋势的研究应当考虑这一因素(Wright, 2002)。
3.4 物种间的相互关系与负密度制约
负密度制约的研究大部分强调同一物种密度及其和相关指标的关系, 聚焦目标物种的种群结构的统计参数变化(Martínez-Ramos et al., 1988; Condit et al., 1994)。在物种丰富的热带森林, 最丰富或者聚集程度最高的物种, 其最近邻体很少是同种个体。因此, 结合异种效应研究物种多样性维持机制至关重要 (Hubbell & Foster, 1986a; Kochummen et al., 1990)。目前引入异种效应对目标物种种群变化的影响来研究负密度制约主要有两种方法: (1)把目标物种周围个体分为同种和异种的邻体, 分析不同类型邻体效应对目标物种种群结构的影响; (2)群落结构引入谱系关系, 分析邻体亲缘关系远近同目标物种动态变化的关系。异种邻体模型考虑了异种邻体与媒介生物之间的相互作用 (Peters, 2003)。
3.4.1 分析异种邻体物种的影响
把目标物种周围的个体分为同种和异种两类, 将异种或同种邻体的个体密度(或者距离)作为单独因子, 分析其与目标物种密度变化(或者个体之间距离的变化)的关系来研究负密度制约(Connell et al., 1984; Uriarte et al., 2004)。有研究表明, 幼苗生长在同种聚集地存活率低, 而多物种混合处的幼苗存活率高(Connell, 1971; Connell et al., 1984)。然而, Wills等(1997)在热带森林BCI样地里的研究却发现, 树木存活率与异种邻体密度没有明显的正相关。
在巴拿马BCI和马来西亚Pasoh热带森林, Peters (2003)调查了同种和异种邻体的聚集格局对树木死亡率的影响, 发现超过80%的物种表现出了同种聚集提高目标个体死亡率的效应, 而异种邻体聚集不会提高目标个体死亡率。于是, Peters(2003)提出物种异群保护模型, 即异种邻体能够阻碍寄主专一的有害生物的传播, 目标个体周围异种邻体的增加, 降低了宿主与其专一性有害生物相遇的机率, 能够提高目标个体的存活率。
3.4.2 引入谱系关系
有研究表明遗传多样性保护可能在物种水平上发挥作用, 从而维持热带森林的物种多样性 (Wills & Green, 1995; Wills, 1996a, b)。因此, 简单地把目标个体周围的物种分成同种和异种两类, 会模糊不同物种对目标物种影响的巨大差异(Pacala et al., 1996)。Uriarte等(2004)在BCI样地对邻体制约幼树生长的研究中发现: 如果目标个体与周围邻体同属于一个科, 那么周围邻体会对目标个体产生更强的负效应。有研究发现两个物种感染同一病原菌的机率与它们之间的系统发育距离呈负相关(Gilbert & Webb, 2007)。
在婆罗洲热带森林, Webb等(2006)用系统发育距离来衡量有机体之间的关系, 局部群落组合引入净关联指数(NRI, net relatedness index)和最近分类指数(NTI, nearest taxon index)来研究幼苗死亡率与其邻体之间的谱系多样性关系。结果发现, 如果幼苗周围邻体谱系多样性提高, 目标物种幼苗的死亡率将会降低。同时基于对生态学和进化生物学的长期研究积累, Webb等(2006)提出了谱系多样性模型。一般来讲, 自然群落中一种病原体感染一组专一宿主的机率会随着宿主之间亲缘关系的距离增大而下降, 也会随着病原体专一性程度的降低而下降。近缘物种比亲缘关系远的物种之间存在更大的负密度制约效应, 因此谱系多样性对维持物种多样性具有一定的贡献。
Webb等(2006)分析了不同尺度上(一个样方大小为1.6 ha, 另一个为36 m2)谱系多样性随着生长阶段的变化, 发现邻体谱系多样性的提高能够降低目标个体幼苗的死亡率。然而, 负密度制约效应受空间尺度的影响, 同一尺度上的结果比较才能比较客观地反映其影响, 而Webb等(2006)的比较是基于两个尺度的, 因此还不能充分证明近缘物种负密度制约效应的存在。
4 研究中存在的主要问题与今后的研究方向
4.1 区域研究不均衡
自1980年以来, 美国史密桑研究院热带森林研究所热带科学研究中心(Center for Tropical Research Smithsonian Tropical Research Institute, CTFS)在15个国家和地区陆续建立了20个大尺度森林动态监测样地, 其大量翔实的数据为检验负密度制约假说提供了便利。然而这样也造成负密度制约的研究主要在热带地区开展, 其他区域的相关研究较少(Hyatt et al., 2003)。因此符合负密度制约假说的物种也主要集中在热带地区(Hille Ris Lambers et al., 2002, 图2)。Carson等(2008)综合分析了53项有关负密度制约的检验, 其中81.2%的研究在热带森林, 15%在温带森林, 只有3.8%的研究在亚热带森林。53个研究总计有7项研究探讨了负密度制约对物种多样性的影响, 其中3项来自BCI, 2项来自BCI和Pasoh, 也就是说负密度制约在群落内普遍存在的检验主要来自BCI样地, 这样就无法探讨负密度制约的发生和强度在不同纬度带的差别。负密度制约是否是跨越不同区域群落的普遍调节机制, 目前这个问题还没有得到明确回答。同时, 在不同纬度带森林群落里, 负密度制约效应发生的特点和强度也有待进一步探讨。
图2
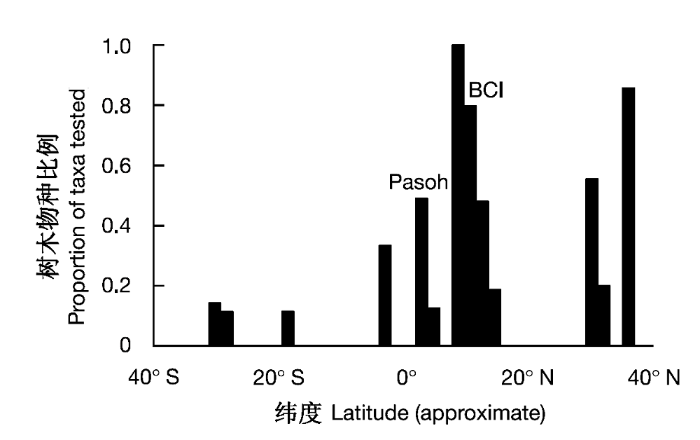
图2
在不同纬度带表现密度制约效应的树木物种比例(Hille Ris Lambers et al., 2002)
Fig. 2
The proportion of tree species showing density dependent effects at different latitudes (Hille Ris Lambers et al., 2002)
中国是生物多样性特别丰富的少数国家之一, 也是唯一一个具有完整气候带谱(从北部的寒温带到南部的热带)的国家, 植被类型丰富。中国科学院生物多样性委员会于2004年启动中国森林生物多样性监测网络(CForBio, Chinese Forest Biodiversity Monitoring Network)建设项目, 已经建成从寒温带到热带4个大于20 ha的样地以及若干5 ha样地(
4.2 负密度制约效应的检验受到其他因素的干扰
众多研究都指出了负密度制约在维持热带森林生物多样性中的重要性。在巴拿马BCI 50 ha热带森林样地中, 有关幼苗和从种子到幼苗早期过渡阶段的研究发现, 负密度制约更新普遍地提高了物种多样性(Harms et al., 2000)。在热带雨林, 不论是理论模型还是观测实验, 都验证了负密度制约能够减少种间竞争排斥, 是物种共存的重要调控机制(Wright, 2002)。然而大部分的野外实验没有排除其他因素的干扰, 比如生境异质性的影响。大尺度的环境异质性能够干扰种子扩散和其他的种群统计参数的准确性, 种群动态和调节机制以一种复杂的方式相互作用, 使空间格局分析变得复杂(Clark et al., 1998; Dieckmann et al., 1999; Condit et al., 2000)。因而生境异质性将会干扰负密度制约效应的检测 (He & Duncan, 2000; Wright, 2002; Getzin et al., 2008)。Zhu等(2009)通过案例—对照分析方法(case-control analysis), 用其他生长阶段的树木分布格局与成年树木格局相比来排除生境异质性效应干扰, 发现了负密度制约效应在中国亚热带古田山24 ha森林存在的普遍性。然而, Zhu等(2009)由于用格局分析来检验负密度制约, 仍需由控制实验来验证。另外主观认为成年树不受负密度制约效应的影响, 或者检验时较少涉及成年树, 也会低估负密度制约的作用。有研究发现成年树也有可能受到负密度制约的影响(Stoll & Newberry, 2005)。未来的研究中需要关注负密度制约效应在成年树阶段的发生与机制。而且, 由于分析方法的限制, 很多研究忽视了稀有种的研究(如Peters 2003; Zhu et al., 2009), 这样就不能客观地理解稀有种和常见种受负密度制约影响的差异。
4.3 距离制约效应的普遍性需要进一步证实
许多野外实验证实了特定物种表现出Janzen-Connell假说的距离制约效应(Augspurger, 1983a; Augspurger & Kelly, 1984; Burkey, 1994; Condit et al., 1994), 然而, 在大尺度的热带森林样地研究中, 仅有少数几个最丰富的物种表现出距离制约效应(Hubbell & Foster, 1986a; Condit et al., 1992; Wills et al., 1997)。个别物种的验证, 难以说明距离制约效应在热带森林群落发生的普遍性, 距离制约效应是否是促进物种共存的重要机制仍受到质疑, 这方面仍需进一步研究。
4.4 格局分析与控制实验
研究者一般经过野外控制实验或者几次样地调查后, 通过分析种群个体死亡率、生长量等的变化来检验负密度制约假说。然而, 目前很难进行大尺度的、长期的实验性研究。在较短的时间间隔内难以检测出负密度制约的信号, 种群空间上的组织可以更好地解释个体死亡的分布格局(Kenkel et al., 1989), 不同生活史阶段个体之间的空间结构不但能够表达长期积累的信号, 推演过去发生的过程, 而且能反过来用于发掘空间结构变化的驱动机制。遗憾的是, 通过格局分析推导实际过程, 两者通常不能一一对应, 因此所提出的理论有可能是观察到的格局的部分发生机制。在未来负密度制约的研究中, 格局分析和控制实验相结合可能有助于更好地解决负密度制约的相关问题。
参考文献
Density dependence and patch dynamics in tropical rain forests: matrix models and applications to a tree species
Demographic and genetic models in conservation biology: applications and perspe- ctives for tropical rain forest tree species
Offspring recruitment around tropical trees: changes in cohort distance with time
Seed dispersal of the tropical tree, Platypodium elegans, and the escape of its seedlings from fungal pathogens
Seedling survival of tropical tree species: interactions of dispersal distance, light gaps, and pathogens
Pathogen mortality of tropical tree seedlings: experimental studies of the effects of dispersal distance, seedling density, and light conditions
DOI:10.1007/BF00396763
URL
PMID:28309414
[本文引用: 1]

We present results of two experiments designed to identify the relative importance of dispersal distance, seedling density, and light conditions on pathogen-caused mortality of tropical tree seedlings. The field experiment on Barro Colorado Island, Panama, demonstrated that both an increase in dispersal distance and a decrease in seedling density reduce levels of damping-off disease among seedlings of Platypodium elegans, and that there is an interaction between the two factors. The results indicated significant variation among sites in pathogen activity and suggested that seedlings are more vulnerable to disease when establishing around their parent tree than around other conspecific trees.The second experiment in a screened enclosure used potted seedlings of 18 wind-dispersed tree species exposed to two levels of sunlight and seedling density. The results indicated that environmental conditions similar to those in light-gaps significantly reduce pathogen activity. They also confirmed that high seedling density increases disease levels, especially under shaded conditions.Seedlings of 16 of the 18 species experienced pathogencaused mortality, but in widely varying amounts. Seed weight was not a good predictor of a species' vulnerability to pathogens. Adult wood density, an indicator of growth rate and successional status, was inversely correlated with a species' vulnerability to pathogens. Fast-growing, colonizing species, whose seedlings require light-gaps, lacked strong resistance to seedling pathogens, relative to slow-growing species able to tolerate shade and escape seedling pathogens. We discuss these results in the context of seed dispersal as a means of escaping from seedling pathogens.
Plant pathogens drive density-dependent seedling mortality in a tropical tree
Some attempts to estimate species diversity and population density of trees in Amazonian forests
Do dipterocarp seedlings really partition tropical rain forest gaps?
Tropical tree species diversity: a test of the Janzen-Connell model
DOI:10.1007/BF00325893
URL
PMID:28313744
[本文引用: 1]
To test the premises and predictions of the Janzen-Connell model (Janzen's spacing mechanism), seeds of the rainforest canopy tree, Brosimum alicastrum, were placed at different distances from the parent tree and their removal observed over 3 weeks. The number and density of naturally occurring seeds at different distances from the parent tree were also estimated. Predation was not greater near the parent tree, except on the very small spatial scale: the proportion of experimental seeds removed was greater 1 m from the trunk than it was 5-25 m from the trunk. Predation was negatively correlated with seed density, not positively as the Janzen-Connell model assumes-presumably due to predator satiation. The density of seeds after predation peaked 5 m from the tree trunk, but this is well within the crown radius of the parent tree. There is a peak in the number of potential recruits at a distance of 10 m from the parent tree, due to the peaked initial distribution of seeds. This peak is caused by the interaction between the seed density curve and the increasing area of an annulus around the parent tree at increasing distances, not by the product of the density curve and the predation curve. However, it is important to realize that it is not the presence of a peak in recruitment away from the parent that is essential to maintaining tropical tree species diversity, but frequency-dependent recruitment induced by poor recruitment near conspecifics. Predator satiation seems to be an important factor in the survival of B. alicastrum seeds, possibly at several spatial scales. The number of seeds produced by the tree is negatively correlated with the loss to predators, and trees that have a fruiting conspecific nearby also suffer lower levels of predation. Seed predation increases as one moves from the forest edge into the interior, creating an edge effect that may have long-term effects on the forest composition and tree species diversity. More studies are needed, for other species, other localities, and larger spatial and temporal scales, on both the Janzen-Connell mechanism and this edge effect.
Spacing dynamics of a tropical forest tree: evaluation of the Janzen-Connell model
Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forest
Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank
Research in large, long-term tropical forest plots
Spatial patterns in the distribution of tropical tree species
Recruitment near conspecific adults and the maintenance of tree and shrub diversity in a neotropical forest
Density-dependence in two understory tree species in a neotropical forest
Compensatory recruitment, growth, and mortality as factors maintaining rain forest tree diversity
Tropical rainforest gaps and tree species diversity
The evolutionary ecology of dispersal
Heterogeneity influences spatial patterns and demographics in forest stands
A canker disease of seedlings and saplings of Tetragastris panamensis (Burseraceae) caused by Botryosphaeria dothidea in a lowland tropical forest
Effects of seedling size, El Niño drought, seedling density, and distance to nearest conspecific adult on 6-year survival of Ocotea whitei seedlings in Panama
DOI:10.1007/s004420000616
URL
PMID:28547488
[本文引用: 1]
We present an analysis of the long-term survival of two cohorts of seedlings of the tropical canopy tree Ocotea whitei (Lauraceae) on a 1-ha plot of mature, lowland moist forest on Barro Colorado Island, Panama. In 1980, we counted an even-aged cohort of seedlings that germinated in 1979, then measured and tagged survivors in 1981. We also measured and tagged a second, smaller cohort of seedlings that germinated in 1981. We followed the subsequent survival of all seedlings through 1985. Seedling mortality was phenotypically, temporally, and spatially non-random. Important correlates of non-random mortality included: (1) seedling size and age, (2) an El Nino drought, and (3) biotic neighborhood. Larger and older seedlings survived better than smaller and younger seedlings, respectively, and the El Nino-related drought of 1982-1983 was associated with elevated mortality rates. Seedling density, which was strongly correlated with the proximity to the nearest conspecific adult, increased mortality. The observed mortality patterns suggest that processes consistent with the Janzen-Connell hypothesis operate during the recruitment phase of O. whitei population dynamics. However, the processes causing the observed density- and distance-dependent mortality may vary with factors such as total seed number, seedling size, and climatic variation, making it difficult to determine whether time-integrated seedling-to-adult spacing mechanisms other than self-thinning operate on a given plant population. After 6 years in the hectare studied, survivors remained densest and most numerous underneath the adult trees. We conclude that only long-term demographic data, collected at a variety of scales on a variety of species, will ultimately answer the question: do Janzen-Connell effects contribute substantially to structuring tropical forests?
Density and distance-to-adult effects of a canker disease of trees in a moist tropical forest
DOI:10.1007/BF00326095
URL
PMID:28312801
[本文引用: 2]
We compared the spatial distribution of stem cankers on the canopy tree Ocotea whitei (Lauraceae) in a 20-ha plot on Barro Colorado Island, Panama, with spatial and temporal patterns of mortality in this host over the previous decade. The cankers occur both on adult and juvenile individuals, aothough juveniles are much more likely the adults to show symptoms. Disease incidence is host-density dependent, and both the presence of the disease and host mortality are more likely close to than far from a conspecific adult, which resulted in a net spatial shift of the juvenile population away from conspecific adults through time. Disease incidence is lower than expected among juveniles of O. whitei growing near to adults of the non-susceptible canopy tree Beilschmiedia pendula. The coincidence of spatial patterns of canker incidence and host mortality suggest a role for the disease in regulating host spatial distribution, in agreement with predictions of the Janzen-Connell hypothesis.
Phylogenetic signal in plant pathogen-host range
The maintenance of species richness in plant communities: the importance of the regeneration niche
Testing the dispersion of juveniles relative to adults: a new analytic method
Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest
Density-dependent effects on tree survival in an old-growth douglas fir forest
Distribution patterns of tree species in a Malaysian tropical rain forest
Density-dependent mortality and the latitudinal gradient in species diversity
DOI:10.1038/nature00809
URL
PMID:12066182
[本文引用: 2]
Ecologists have long postulated that density-dependent mortality maintains high tree diversity in the tropics. If species experience greater mortality when abundant, then more rare species can persist. Agents of density-dependent mortality (such as host-specific predators, and pathogens) may be more prevalent or have stronger effects in tropical forests, because they are not limited by climatic factors. If so, decreasing density-dependent mortality with increasing latitude could partially explain the observed latitudinal gradient in tree diversity. This hypothesis has never been tested with latitudinal data. Here we show that several temperate tree species experience density-dependent mortality between seed dispersal and seedling establishment. The proportion of species affected is equivalent to that in tropical forests, failing to support the hypothesis that this mechanism is more prevalent at tropical latitudes. We further show that density-dependent mortality is misinterpreted in previous studies. Our results and evidence from other studies suggest that density-dependent mortality is important in many forests. Thus, unless the strength of density-dependent mortality varies with latitude, this mechanism is not likely to explain the high diversity of tropical forests.
On mechanisms of species coexistence in plant communities
Survival and growth of juvenile Virola surinamensis in Panama: effects of herbivory and canopy closure
Ecology of seed dispersal
Tree dispersion, abundance, and diversity in a tropical dry forest
Seed predation and the coexistence of tree species in tropical forests
A unified theory of biogeography and relative species abundance and its application to tropical rain forests and coral reefs
Local neighborhood effects on long-term survival of individual trees in a neotropical forest
Presence and absence of density dependence in a neotropical tree community
The distance dependence prediction of the Janzen-Connell hypothesis: a meta-analysis
Herbivores and the number of tree species in tropical forests
Local competition in a naturally established jack pine stand
Floristic composition of the Pasoh Forest Reserve, a lowland rain forest in Peninsular Malaysia
Why do some tropical forests have so many species of trees?
Distance-dependent effects of soil-derived biota on seedling survival of the tropical tree legume Ormosia semicastrata
Large scale permanent plots: important platform for long term research on biodiversity in forest ecosystem
Tolerance of soil pathogens co-varies with shade tolerance across species of tropical tree seedlings
DOI:10.1890/07-0211.1
URL
PMID:18705375
[本文引用: 1]
A negative feedback between local abundance and natural enemies could contribute to maintaining tree species diversity by constraining population growth of common species. Soil pathogens could be an important mechanism of such noncompetitive distance and density-dependent (NCDD) mortality, but susceptibility to local pathogens may be ameliorated by a life history strategy that favors survivorship. In a shade-house experiment (1% full sun), we tested seedling life span, growth, and mass allocation responses to microbial extract filtered from conspecific-cultured soil in 21 tree species that varied in abundance and shade tolerance in a wet tropical forest (La Selva Biological Station, Costa Rica). Forty-three percent of the species had significant reductions, and 10% of the species had significant increases in life span, growth, root length, or root surface area when inoculated with microbial extract; 10% of the species experienced opposing reductions and increases in these characteristics. Contrary to expectation, species' local abundance was not related to species-specific responses to microbial extracts from cultured soils. Across species, seedling shade tolerance (survival at 1% full sun) was negatively correlated with susceptibility to the microbial. treatment for both survival and total mass accumulation, thereby exaggerating shade tolerance differences among species. Thus, soil pathogens may contribute to species coexistence through heightening niche differentiation rather than through negative density dependence in common species.
Forest models defined by field measurements: estimation, error analysis and dynamics
Soil pathogens and spatial patterns of seedling mortality in a temperate tree
Janzen-Connell effects are widespread and strong enough to maintain diversity in grasslands
DOI:10.1890/07-2056.1
URL
PMID:18831160
[本文引用: 1]
Crop rotation schemes are believed to work by preventing specialist soil-borne pests from depressing the future yields of similar crops. In ecology, such negative plant-soil feedbacks may be viewed as a type of Janzen-Connell effect, which promotes species coexistence and diversity by preventing the same species from repeatedly occupying a particular site. In a controlled greenhouse experiment with 24 plant species and using soils from established field monocultures, we reveal community-wide soil-based Janzen-Connell effects between the three major functional groups of plants in temperate European grasslands. The effects are much stronger and more prevalent if plants are grown in interspecific competition. Using several soil treatments (gamma irradiation, activated carbon, fungicide, fertilizer) we show that the mechanism of the negative feedback is the buildup of soil pathogens which reduce the competitive ability of nearly all species when grown on soils they have formerly occupied. We further show that the magnitude of the change in competitive outcome is sufficient to stabilize observed fitness differences between functional groups in reasonably large communities. The generality and strength of this negative feedback suggests that Janzen-Connell effects have been underestimated as drivers of plant diversity in temperate ecosystems.
Neighbour-regulated mortality: the influence of positive and negative density dependence on tree populations in species-rich tropical forests
An estimate of the number of species of trees in an Amazonian forest community
Neighborhood and community interactions determine the spatial pattern of tropical tree seedling survival
DOI:10.1890/06-0737.1
URL
PMID:17918403
[本文引用: 1]
Factors affecting survival and recruitment of 3531 individually mapped seedlings of Myristicaceae were examined over three years in a highly diverse neotropical rain forest, at spatial scales of 1-9 m and 25 ha. We found convincing evidence of a community compensatory trend (CCT) in seedling survival (i.e., more abundant species had higher seedling mortality at the 25-ha scale), which suggests that density-dependent mortality may contribute to the spatial dynamics of seedling recruitment. Unlike previous studies, we demonstrate that the CCT was not caused by differences in microhabitat preferences or life history strategy among the study species. In local neighborhood analyses, the spatial autocorrelation of seedling survival was important at small spatial scales (1-5 m) but decayed rapidly with increasing distance. Relative seedling height had the greatest effect on seedling survival. Conspecific seedling density had a more negative effect on survival than heterospecific seedling density and was stronger and extended farther in rare species than in common species. Taken together, the CCT and neighborhood analyses suggest that seedling mortality is coupled more strongly to the landscape-scale abundance of conspecific large trees in common species and the local density of conspecific seedlings in rare species. We conclude that negative density dependence could promote species coexistence in this rain forest community but that the scale dependence of interactions differs between rare and common species.
The dispersal of plants throughout the world
Seed and early seedling predation in the forest understory and in treefall gaps
The Janzen-Connell model for tropical tree diversity: population implication and the importance of spatial scale
The role of density dependence in the population dynamics of a tropical palm
Evidence of species-specific neighborhood effects in the Dipterocarpaceae of a Bornean rain forest
On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies
A spatially explicit model of sapling growth in a tropical forest: does the identity of neighbours matter?
Density dependence explains tree species abundance and diversity in tropical forests
DOI:10.1038/nature04030
URL
PMID:16319890
[本文引用: 1]
The recurrent patterns in the commonness and rarity of species in ecological communities--the relative species abundance--have puzzled ecologists for more than half a century. Here we show that the framework of the current neutral theory in ecology can easily be generalized to incorporate symmetric density dependence. We can calculate precisely the strength of the rare-species advantage that is needed to explain a given RSA distribution. Previously, we demonstrated that a mechanism of dispersal limitation also fits RSA data well. Here we compare fits of the dispersal and density-dependence mechanisms for empirical RSA data on tree species in six New and Old World tropical forests and show that both mechanisms offer sufficient and independent explanations. We suggest that RSA data cannot by themselves be used to discriminate among these explanations of RSA patterns--empirical studies will be required to determine whether RSA patterns are due to one or the other mechanism, or to some combination of both.
Phylodiversity- dependent seedling mortality, size structure, and disease in a Bornean rain forest
Seedling density dependence promotes coexistence of Bornean rain forest trees
Sapling survival, growth, and recruitment: relationship to canopy height in a neotropical forest
Similar non-random processes maintain diversity in two tropical rainforests
DOI:10.1098/rspb.1999.0799
URL
PMID:10457617
[本文引用: 1]
Quadrat-based analysis of two rainforest plots of area 50 ha, one in Panama (Barro Colorado Island, BCI) and the other in Malaysia (Pasoh), shows that in both plots recruitment is in general negatively correlated with both numbers and biomass of adult trees of the same species in the same quadrat. At BCI, this effect is not significantly influenced by treefall gaps. In both plots, recruitment of individual species is negatively correlated with the numbers of trees of all species in the quadrats, but not with overall biomass. These observations suggest, but do not prove, widespread frequency-dependent effects produced by pathogens and seed-predators that act most effectively in quadrats crowded with trees. Within-species correlations of mortality with numbers or biomass are not found in either plot, indicating that most frequency-dependent mortality takes place before the trees reach 1 cm in diameter. Stochastic effects caused by BCI's more rapid tree turnover may contribute to a larger variance in diversity from quadrat to quadrat at BCI, although they are not sufficient to explain why BCI has fewer than half as many tree species as Pasoh. Finally, in both plots quadrats with low diversity show a significant increase in diversity over time, and this increase is stronger at BCI. This process, like the frequency-dependence, will tend to maintain diversity over time. In general, these non-random forces that should lead to the maintenance of diversity are slightly stronger at BCI, even though the BCI plot is less diverse than the Pasoh plot.
Strong density- and diversity-related effects help to maintain tree species diversity in a neotropical forest
A genetic herd-immunity model for the maintenance of MHC polymorphism
Nonrandom processes maintain diversity in tropical forests
Plant diversity in tropical forests: a review of mechanisms of species coexistence
DOI:10.1007/s004420100809
URL
PMID:28547014
[本文引用: 13]
Evidence concerning mechanisms hypothesized to explain species coexistence in hyper-diverse communities is reviewed for tropical forest plants. Three hypotheses receive strong support. Niche differences are evident from non-random spatial distributions along micro-topographic gradients and from a survivorship-growth tradeoff during regeneration. Host-specific pests reduce recruitment near reproductive adults (the Janzen-Connell effect), and, negative density dependence occurs over larger spatial scales among the more abundant species and may regulate their populations. A fourth hypothesis, that suppressed understory plants rarely come into competition with one another, has not been considered before and has profound implications for species coexistence. These hypotheses are mutually compatible. Infrequent competition among suppressed understory plants, niche differences, and Janzen-Connell effects may facilitate the coexistence of the many rare plant species found in tropical forests while negative density dependence regulates the few most successful and abundant species.
Density dependence is prevalent in a heterogeneous subtropical forest
DOI:10.1111/j.2007.0030-1299.15372.x
URL
PMID:32367896
[本文引用: 5]
Environmental change is not likely to act on biodiversity in a random manner, but rather according to species traits that affect assembly processes, thus, having potentially serious consequences on ecological functions. We investigated the effects of anthropogenic land use on functional richness of local hoverfly communities of 24 agricultural landscapes across temperate Europe. A multivariate ordination separated seven functional groups based on resource use, niche characteristics and response type. Intensive land use reduced functional richness, but each functional group responded in a unique way. Species richness of generalist groups was nearly unaffected. Local habitat quality mainly affected specialist groups, while land use affected intermediate groups of rather common species. We infer that high species richness within functional groups alone is no guarantee for maintaining functional richness. Thus, it is not species richness per se that improves insurance of functional diversity against environmental pressures but the degree of dissimilarity within each functional group.

{kind=link}
{kind=link}
{kind=link}
{kind=link}