


植物群落是指生活在一定区域内所有植物的集合, 它是每个植物个体通过互惠、竞争等相互作用而形成的一个巧妙组合, 是适应其共同生存环境的结果。在人类文明进步的历史进程中, 植物群落提供了人类赖以生存的主要物质资源, 具有不可替代性。植物群落的重要性主要在于:
(1) 植物群落是物种的载体, 汇聚了各类生物资源: 植物群落是不同植物在长期环境变化中相互适应而形成的, 它聚集了各类野生植物品种资源(如野生稻、野生大豆等)、中草药以及珍稀濒危植物, 也为各种动物和其他生物提供着食物来源以及栖息地。因此, 植物群落不仅为人类提供赖以生存的种质资源, 也是利用、开发和保护其他生物资源的基础。
(2) 植物群落是提供生态系统功能的主体: 植物生物量占全球总生物量的99%(Lieth, 1975), 是生态系统的生产者。植物群落还具备其他重要的生态功能, 如吸收大气中的CO2, 减缓温室效应, 控制水土流失, 减轻水体和大气污染等等。植物群落在维持和改善人类生存环境方面具有不可替代的作用。
(3) 植物群落是土地基本属性的综合指标: 特定的气候、土壤和地形条件发育了不同的植物群落, 植物群落则综合反映了土地的基本属性。因此, 植物群落的整体状况综合体现了国家的生态本底, 是生态恢复和生态建设以及制定土地利用政策的重要依据。
充分认识植物群落的重要性和它的价值是开发、利用和保护生物资源的前提, 也是生态学、环境科学和地理学等相关学科发展的需要。为此,我们有必要对其进行全面系统的清查。植物群落清查(plant community inventory)就是用统一、规范的调查方法, 对植物群落进行全面、系统的野外调查, 在此基础上, 掌握植物群落的整体现状(包括群落类型及其物种构成、结构、分布和动态等), 分析群落与环境的相互关系, 对重点群落类型进行长期监测, 了解群落优势种的生态属性等等, 并对群落现状和发展趋势进行评估。群落清查的主要成果是群落数据库和群落图志, 并为生物多样性利用和保护、土地利用状况的监测、生态系统管理、区域发展规划等提供基础资料。
世界上发达国家都无一例外地从国土生态安全和生物资源可持续利用的战略高度, 重视植物群落的调查工作。目前最有影响的国家层面的植物群落调查计划, 包括欧洲的“植被测量组织”(European Vegetation Survey, EVS)(Schaminée et al., 2009)、美国的“植被库计划”(VegBank) (Bekker et al., 2007), 以及日本的“国土绿色调查”(緑の国勢調査) (
我国地域广袤, 环境梯度大, 跨越从寒温带至热带、雨林到荒漠、海岛到高原的自然环境, 生态系统类型复杂多样。因此, 我国植物群落类型的环境指示性和类型多样性在世界上首屈一指, 是一笔价值无量的国家财富。尽管我国已于20世纪50-90年代对西藏、横断山区、新疆、内蒙、宁夏、陕甘黄土高原等地进行过大规模的植被考察工作, 也出版了一批重要的植被学专著或植被图, 如《中国植被》(吴征镒, 1980)和《1:100万中国植被图图集》及其说明书(侯学煜, 2001; 张新时, 2007), 但这些工作基本上都是基于植被考察的成果, 实际上我国至今仍没有进行过一次全面和系统的植物群落清查。我国到底有多少种植物群落类型?其组成和分布如何?植物群落对区域气候的指示作用和对气候变化的响应怎样?人类干扰对植物群落的演化产生怎样的影响?我国生物资源与区域经济开发的关系如何?等等诸多的科学问题都亟待我们回答。全面系统地植物群落清查将为回答这些问题提供不可或缺的基础数据。
基于前人的工作积累, 结合作者们长期的野外工作实践, 本文从样方调查、生境(地形、气候、土壤)调查、群落复查(监测) 以及植物生态属性测定等方面, 总结和归纳出一套较为系统的群落清查方法体系和技术规范, 试图为我国开展群落清查工作提供技术储备。本文提供的一些群落调查方法也可作为一般群落调查的参考。
在论述群落调查方法之前, 有必要先对群落清查中的两个重要概念“样地”和“样方”进行定义。样地和样方是两个既关联又有区别的空间概念。在本文中, 样地(site)指群落调查的所在地, 在空间上它包含样方, 一般没有特定的面积; 而样方(plot)则指群落调查所要实施的特定地段, 有特定的面积, 如森林调查的样方一般为600 m2。
1 群落清查中的重要概念和测度
1.1 种–面积曲线与巢式取样
种–面积曲线 (species-area curve)或种–面积关系 (species-area relationship)是群落调查的重要内容之一。原则上, 调查样方的面积大小是根据种–面积曲线确定的。一般来说, 物种越丰富的群落, 设置的样方面积也应越大。
种–面积曲线是非常重要的群落特征, 它描述了物种数量随面积增加而增加的规律, 其机制在于: (1) 取样面积的增加可以包含更多的生境异质性, 因此可包含更多的物种数; (2) 随着取样面积的增加, 所包含的个体数也将增加, 从而具有包含更多物种的可能; (3)某些进化或生态过程仅发生在面积足够大的生境。关于种–面积曲线的内涵、机制和意义等, 详见唐志尧等(2009)。
描述种–面积曲线的主要模型有3种(Gurevitch et al., 2002):
上式中, S为物种数, A为样方面积, b, c和z为常数。
在这3种模型中, 最常用的是Arrhenius幂函数模型。通常情况下, 它可以很好地拟合实际的种–面积关系。该模型的主要优点在于幂指数(z)的意义很明确, 数值的变化范围较小, 一般变动于0.15- 0.50之间, 并且呈现由热带向寒带递减的趋势(Drakare et al., 2006)。Wang等(2009)对东亚和北 美植物多样性的大尺度分析证明了这种趋势, 并 且发现在相同热量条件下, 东亚地区的z值高于 北美。
图1
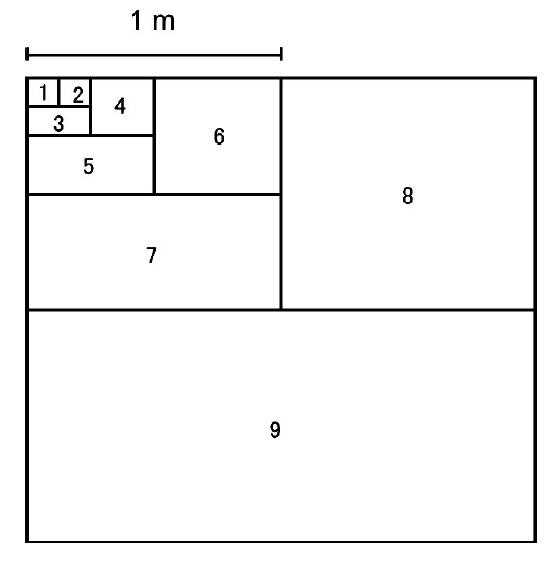
图1
巢式取样示意图。数值表示取样顺序。
Fig. 1
Illustration of nested sampling. The numbers indicate the sampling sequence.
1.2 物种重要性的直接测度
植物群落是由不同植物物种组成的。一种植物在群落中的重要性如何, 可由多个指标来量度。通过这些指标的测量, 回答该物种是否存在、数量多少、个体多大等问题。群落调查中直接测定的物种重要性测度常常包括: 出现/不出现、盖度(郁闭度)、植株密度、多度、直径和高度等。
(1) 出现/不出现(presence or absence): 指某种植物在样方中是否存在, 以该植物个体的基部是否生长在所调查的样方中为准。换言之, 地上部分出现在样方中但其基部并不生长在样方内的植株不能计入该样方。
(2) 盖度(coverage): 指植物地上部分垂直投影面积占样方面积的百分比, 又称投影盖度。群落调查时, 可以记载每个优势种的盖度(称种盖度或分盖度), 种盖度之和可以超过100%, 但任何单一种的盖度都不会大于100%。
为计测方便起见, 常常将物种的盖度划分为若干个盖度级。本文推荐使用Braun-Blanquet(1964)的盖度级分级标准(表1)。
表1 Braun-Blanquet的盖度分级标准
Table 1
| 分级 Scale | + | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|---|
| 盖度范围 Range of coverage (%) | 少有出现 Rare | 0-5 | 5-25 | 25-50 | 50-75 | >75 |
对森林群落而言, 常用郁闭度(canopy coverage)来表示乔木层的盖度, 它是指林冠覆盖面积与地表面积之比, 常以十分数表示, 即林冠完全覆盖地面记为1.0。一般来说, 郁闭度≥0.70的为密林, 0.20-0.69为中度郁闭, < 0.20为疏林。
(3) 植株密度(density of individuals): 指样方中的植物个体数量。每种植物有各自的个体数量, 称种群密度(population density)。所有物种的种群密度之和即是群落的个体密度。对于森林而言, 群落的乔木层植株密度也称林分密度(stand density)。
(4) 多度(abundance): 是一种物种个体数量的目测估计指标, 主要用于快速获得盖度的野外调查, 常采用Drude的七级制进行分级(表2)。如果测定了群落的盖度或密度, 则可以不测定多度。
表2 Drude的多度分级标准
Table 2
| 分级 Scale | 7 | 6 | 5 | 4 | 3 | 2 | 1 |
|---|---|---|---|---|---|---|---|
| 符号 Symbol | soc | cop3 | cop2 | cop1 | sp | sol | un |
| 描述 Description | 极多 | 很多 | 多 | 尚多 | 不多 | 稀少 | 单株 |
(5) 胸高直径(简称胸径, diameter at breast height, DBH): 木本植物的茎干直径是森林群落调查中最重要、也最易测定的指标, 常常被用来表示群落的大小。群落分析中常常使用的胸高断面积(basal area)和生物量就是由DBH来推算的。一般来说, 对于树高超过胸高部位(我国及国际上大多数国家取1.3 m处, 美国取1.4 m或4.5 ft)的个体, 测其DBH; 反之, 可测其基部直径(简称基径)。
(6) 树高(tree height): 也是一种非常重要的群落生长因子, 既体现乔木树种的生物学特性和该树种的生长能力, 也是判别群落立地质量的指标, 并指示森林生物量的高低。在全球尺度上, 郁闭森林的地上生物量与树高之比为一常数, 即10.6 t·ha-1·m-1), 也就是说, 单位森林空间的地上生物量密度恒定, 为1.0 kg/m3 (Fang et al., 2006)。这足以说明树高在群落调查中的重要性, 但树高的测定较为困难, 尤其在高大郁闭的森林中。因此, 实践上常常只测定部分个体的树高, 然后通过建立树高与DBH之间的相关生长关系, 由DBH估算树高。
(7) 重要值(importance value, IV): 也是一个重要的群落定量指标, 并常用于比较不同群落间某一物种在群落中的重要性, 它通过上述直接测度指标计算得到, 并非直接测量的。一般计算式为:
其中, 相对多度(%) = 100×某个种的株数/所有种的总株数; 相对频度(%) = 100×某个种在统计样方中出现的次数/所有种出现的总次数; 相对优势度(%) = 100×某个种的胸高断面积/所有种的胸高断面积。
计算式(4)并非是重要值的唯一计算方法, 它可根据群落类型和已有数据作相应的变动。如在草本群落中, 可用物种的平均高度替代优势度, 或相对盖度替代相对多度进行计算; 在森林群落中, 常常直接用乔木层的相对优势度(相对胸高断面积)来表示重要值。总之, 在具体的研究中, 需对重要值的计算进行定义。
1.3 群落多样性及其测度
群落多样性(community diversity)是生物群落的重要特征, 反映群落自身特征及其与环境之间的相互关系。群落多样性一般包括α多样性和β多样性。α多样性表示群落中所含物种的多少, 即物种丰富度(species richness), 以及群落中各个种的相对密度, 即物种均匀度(species evenness)。β多样性则表示物种沿环境梯度所发生替代的程度或物种变化的速率。不同群落或某一环境梯度上不同样方之间的共有种越少, β多样性越大; 反之亦然。另外, 在较大的地理空间上, 常常用γ多样性来指示一个区域内总的物种多样性数量。
表示群落多样性的指标繁多(Whittaker, 1972; Magurran, 1988; 马克平, 1994; Spellerberg & Fedor, 2001; Veech et al., 2002), 建议使用如下指标测度:
(1) 物种丰富度(S)
(2) α多样性(Magurran, 1988)
其中: Pi为种i的相对优势度(相对胸高断面积)或重要值(IV)。
(3) β多样性(Whittaker, 1972; Magurran, 1988)
其中, a和b分别为两样方的物种数, c为两样方的共有物种数, g(H)为沿生境梯度H增加的物种数, l(H)为沿生境梯度H失去的物种数。
上述指数中, Sørensen指数和Jaccard指数反映群落或样方间物种组成的相似性; Cody指数则反映样方物种组成沿环境梯度的替代速率。
2 调查样地的设置原则和体系
为全面掌握一个地区或国家的群落现状、变化及所在地的环境条件, 同时考虑到人力和财力的限制, 群落清查时, 需要对样地的布局进行合理设计。根据中国目前的状况, 建议从系统布点、全面调查和重点精查3个层面开展中国植物群落的清查。实际上, 这3个层面也体现了样地布局的原则, 即全面性、代表性和典型性。
全面性指样地在空间上涵盖整个研究区, 布局均衡, 能够反映研究区植被和环境的全貌; 代表性指布点必须包含所有代表性的植物群落类型, 是群落清查的主体内容; 典型性指布点时应保证研究区内典型和特殊植物群落得到重点和细致的调查, 为群落复查和长期监测服务。
(1) 系统布点: 采用统一的经纬网格, 对研究区的植物群落进行系统布点。这样可以达到全面调查研究区植物群落及其生境的目的。经纬网格的精度可根据任务要求、群落类型的复杂程度以及研究区的面积大小作相应要求, 如0.1˚×0.1˚, 0.2˚×0.2 ˚, 0.5˚×0.5˚等(图2)。每个网格的样地可统一设置在网格的四角或中央, 每个样地设置3-5个重复样方。森林群落的样方面积为20 m×30 m (此值与国内常用的面积单位“亩”, 即667 m2基本一致)。
图2
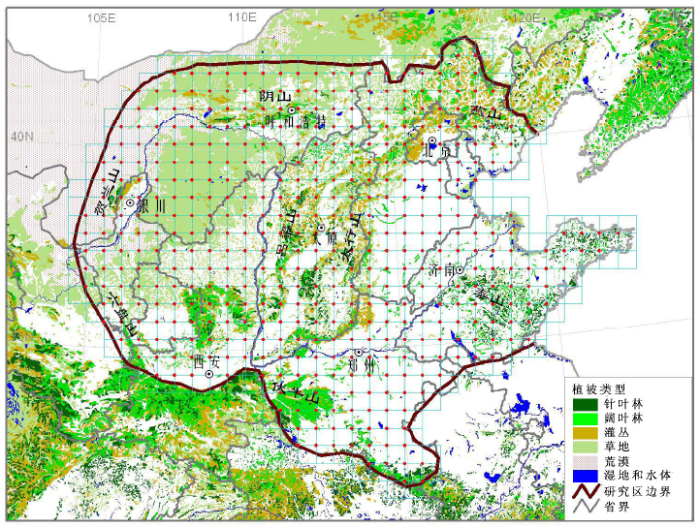
样方调查内容可包括群落调查、环境因子和重要物种生态属性的测定。图2以华北及周边地区的群落清查为例, 显示网格的布局。
(2) 全面调查: 根据1:100万中国植被图和区域群落记载资料, 全面调查研究区的植物群落, 保证研究区中每一种主要自然群落类型都能得到调查; 在山地, 要按海拔高度和植被类型设置样地。也就是说, 样地的多少取决于自然植物群落类型的数量。森林群落的样方面积为20 m×30 m, 每个样地设置3-5个重复样方。
样方调查内容主要包括群落调查以及部分样方的环境因子和重要物种生态属性的测定。在群落调查时, 除一般的测定项目外, 还要在现场手绘植物群落剖面图(图3), 以反映群落的空间结构和种间关系等群落特征。
图3

图3
植物群落剖面示意图。引自《福建植被》 (林鹏, 1990)。调查地点为翠安县三港二里坪, 海拔770 m。1: 甜槠; 2: 青冈; 3: 东南石栎; 4: 光叶石楠; 5: 南岭山矾; 6: 鹿角杜鹃; 7: 马银花; 8: 粗叶木; 9: 肿节竹; 10: 细齿叶柃; 11: 中华里白。
Fig. 3
Illustration of vertical structure of the forest comm- unity (from Lin, 1990). The plot is located at Erliping, Cuian County, Fujian, at an altitude of 770 m above sea level. The species are: 1, Castanopsis eyrei; 2, Cyclobalanopsis glauca; 3, Lithocarpus harlandii; 4, Photinia glabra; 5, Symplocos confusa; 6, Rhododendron latoucheae; 7, R. ovatum; 8, Lasianthus chinensis; 9, Oligostachyum oedogonatum; 10, Eurya nitida; 11, Hicriopteris chinensis.
(3) 重点精查: 对研究区的地带性、特有、稀有、濒危以及有特殊用途和重要经济价值的群落进行精查。精查对象也包括有重要学术价值, 如分布在植被带的南界或北界以及呈隔离状态的植物群落。森林群落的样方面积为20 m×50 m, 每个样地设置3-5个重复样方。
样方调查内容主要包括群落调查、环境因子测定以及重要物种生态属性的测定。在群落调查项目中, 也包括植物群落剖面图的绘制。
3 群落清查的内容、技术指标和方法
根据植被类型及其结构特征的差异, 将植物群落分为森林、灌丛和草地, 以及水生植物群落, 分别规定其清查内容和技术规范。另外, 本文中不讨论荒漠群落的清查, 但可参照灌丛和草地植被的调查体系。由于荒漠植被稀疏且异质性大, 调查面积应大于灌丛和草地的调查面积。
3.1 群落调查表及说明
附表1 森林群落样方基本信息表
Appendix 1
| 样方编号 | 群落类型 | 样方面积 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 调查地点 | 省 县(林业局) 乡(林场) 村(林班) | |||||||||
| 具体位置描述: | ||||||||||
| 纬度 | 地形 | ( )山地 ( )洼地 ( )丘陵 ( )平原 ( )高原 | ||||||||
| 经度 | 坡位 | ( )谷地 ( )下部 ( )中下部 ( )中部 | ||||||||
| 海拔 | ( )中上部 ( )山顶 ( )山脊 | |||||||||
| 坡向 | 森林起源 | ( )原始林 ( )次生林 ( )人工林 | ||||||||
| 坡度 | 干扰程度 | ( )无干扰 ( )轻微 ( )中度 ( )强度 | ||||||||
| 土壤类型 | 林龄 | 群落剖面图: | ||||||||
| 垂直结构 | 层高(m) | 盖度(%) | 优势种 | |||||||
| 乔木层 | ||||||||||
| 亚乔木层 | ||||||||||
| 灌木层 | ||||||||||
| 草本层 | ||||||||||
| 调查人 | ||||||||||
| 记录人 | 调查日期 | |||||||||
附表2 群落调查记录表 Appendix 2 Records for plot survey
附表2-b 灌木层调查表
Appendix 2b
 |
注: 1. 在S1、S2灌木样方中调查时, 记录每丛(株)的种名、平均基径、平均高、株数。
2. 在其他样方中调查时, 仅记录未在S1、S2中出现的物种的种名。
附表2-c 草本层调查表
Appendix 2c
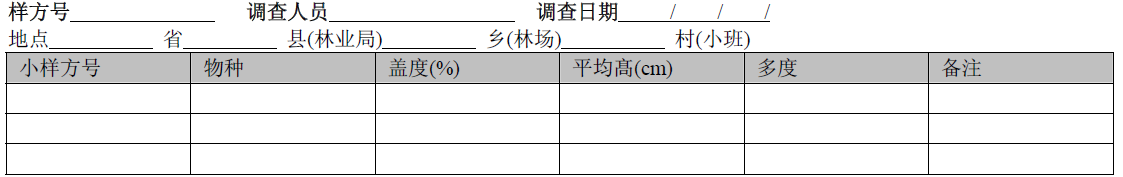 |
注: 1. 在H1–H5草本小样方中调查时, 记录每个种的盖度、平均高、多度。按德氏多度等级记载多度: 极多–soc, 很多–cop3, 多–cop2, 尚多–cop1, 不多–sp, 稀少–sol, 仅1株–un。
2. 在10×10 m样方中调查时, 仅记录未在H1–H5小样方中出现的物种名。
(1) 群落类型: 样方的群落类型。
(2) 调查地: 样方的所在位置, 如区县市村镇或林业局(场)小班和保护区名称, 并标在地形图上。
(3) 经纬度: 用GPS确定样方所在地的经纬度。
(4) 海拔: 用海拔表确定样方所在地的海拔。值得注意的是, GPS测定海拔高度的误差较大, 应尽量避免使用GPS测定海拔高度。
(5) 地形: 样方所在地的地貌类型, 如山地、洼地、丘陵、平原等。
(6) 坡位: 样方所在坡面的位置, 如谷地、下部、中下部、中部、中上部、山顶、山脊等。
(7) 坡向: 样方所在地的方位, 以S30˚E(南偏东30度)的方式记入。
(8) 坡度: 样方的平均坡度。
(9) 面积: 样方的面积, 一般为600 m2或1,000 m2, 记为20 m×30 m或20 m×50 m。
(10) 土壤类型: 样方所在地的土壤类型, 如褐色森林土、山地黄棕壤等。
(11) 森林起源: 按原始林、次生林和人工林 记录。
(12) 干扰程度: 按无干扰、轻微、中度、强度干扰等记录。
(13) 群落层次: 记录群落垂直结构的发育程度, 如乔木层、灌木层、草本层等是否发达等。
(14) 优势种: 记录各层次的优势种, 如某层有多个优势种, 要同时记录。
(15) 群落高度: 群落的大致高度, 可给出范围, 如15-18 m。
(16) 郁闭度: 各层的郁闭度, 用百分比表示。
(17) 群落剖面图: 该图对了解群落的结构、种间关系、地形等非常重要。示例见图3。
(18) 调查人、记录人及日期: 记录该群落的调查人和记录人, 并注明调查日期, 以备查用。
(19) 群落调查记录表: 记录群落的各调查项目, 包括物种、DBH、树高及其他特征, 见附表2。
3.2 森林植物群落清查
森林是植物群落清查的重点。调查样方的面积为20 m×30 m, 观测记录包括乔木层、灌木层、草本层和层间植物。各层次的具体调查内容如下:
(1) 乔木层: 记录样方内出现的全部乔木种, 测量所有DBH≥3 cm的植株胸径和高度, 记录其存活状态。
(2) 灌木层: 记录样方内出现的全部灌木种。选择面积为10 m×10 m的两个对角小样方进行调查, 对其中的全部灌木分种计数, 并测量基径和高度。
(3) 草本层: 记录样方内出现的全部草本种类。测量和记录样方四角和中心点上共5个1 m×1 m的草本层小样方中, 每种草本植物的多度和盖度。
(4) 层间植物: 记录出现的全部寄生、附生植物和攀援植物种类, 并估计其多度和盖度。
具体的调查步骤和技术规范如下。
3.2.1 样方地点的选择
选择适当的地点是样方调查的关键, 在样方选择时应注意: (1) 群落内部的物种组成、群落结构和生境相对均匀; (2) 群落面积足够, 使样方四周能够有10-20 m以上的缓冲区(图4); (3) 除依赖于特定生境的群落外, 一般选择平(台)地或缓坡上相对均一的坡面, 避免坡顶、沟谷或复杂地形。
图4
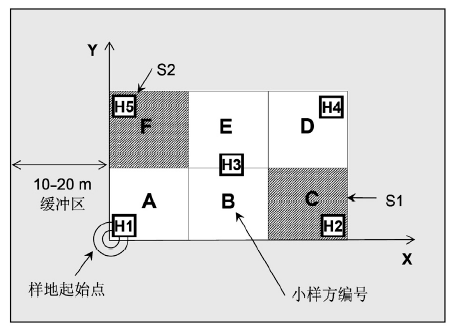
图4
森林群落样方设置和样格编号方法。样方面积20 m×30 m, 由6个10 m×10 m的样格组成, A-F为样格编号, S1和S2(阴影部分)为灌木层调查样格; H1-5为草本调查小样方。样方四边应各留有10-20 m以上的缓冲区。
Fig. 4
Plot setting and quadrate coding for forest communities. The 20 m × 30 m plot is composed of six quadrates (A-F), each with an area of 10 m×10 m. The shadowed quadrates (S1 and S2) were selected for shrub layer investigation, and subplots (H1-5) were selected for herbaceous layer investigation. A buffering zone of 10-20 m at each side of the plot is necessary to keep the plot away from apparent human activities.
3.2.2 样方设置
(1) 样方面积600 m2 (重点精查群落为1,000 m2), 一般为20 m×30 m (20 m×50 m)的长方形。如实际情况不允许, 也可设置为其他形状, 但必须由6 (或10)个10 m×10 m的小样方组成。本文把这种10 m×10 m的小样方称作样格(module)。一般来说, 样方面积有大有小, 但一个样格的面积是固定不变的, 特指10 m×10 m的小样方。
(2) 以罗盘仪确定样方的四边, 闭合误差应在0.5 m以内。以测绳或塑料绳将样方划分为10 m×10 m的样格(图4)。
(3) 对于连续监测样方, 以硬木材质的木桩标记样方的四边和网格, 样方四边木桩地上部分留30 cm左右, 内部网格木桩地上部分留15 cm左右(如条件允许, 可以将磁铁埋在各木桩的位置, 以防人为破坏的影响)。
3.2.3 样方环境因子调查
调查项目详见群落调查表。除调查表所记载的项目外, 还需完成如下项目:
(1) 群落照片: 包括群落外貌、群落垂直结构、乔木层、灌木层、草本层和土壤剖面等。数码照片的分辨率应在300万像素以上。
(2) 温湿度测定: 可采用美国Onset公司的HOBO温湿度自动记录仪测定, 采样频率为10 min, 测定时间1年以上。空气温湿度测定时, HOBO应固定在离地表1-2 m、不会受到阳光直射的树干上; 土壤温湿度测定中, HOBO应埋在距地表10 cm处。
3.2.4 乔木层调查
(1) 记录林分状况: 个体所属层次(乔木层/亚乔木层/更新层)、健康状况(正常/折枝/倾斜/翻倒/濒死/枯立/枯倒)。
(2) 树木编号: 由样格号+树号组成。对于连续监测样方, 每个个体挂上预先统一制作的识 别牌。
(3) 物种记录: 从事群落调查的人员常常会遇到物种分类的困难。因此, 需要采集标本进行鉴定。为便于标本采集和鉴定, 一般要求在野外确认到属。为此, 可提前准备研究区的植物名录以便查对, 并事先进行物种鉴定的培训。
(4) 胸径测定: 在每个样格中, 对于所有DBH≥ 3 cm的树木个体, 记录种名, 测量DBH。对于连续监测样方, 须在DBH测量处进行标记。
DBH是最主要且又易于测定的生长指标, 需要对满足测定标准的每个个体都进行准确测定。对于生长不规则的树木, 测定DBH时, 应注意以下事项(胸径测定位置见图5):
图5
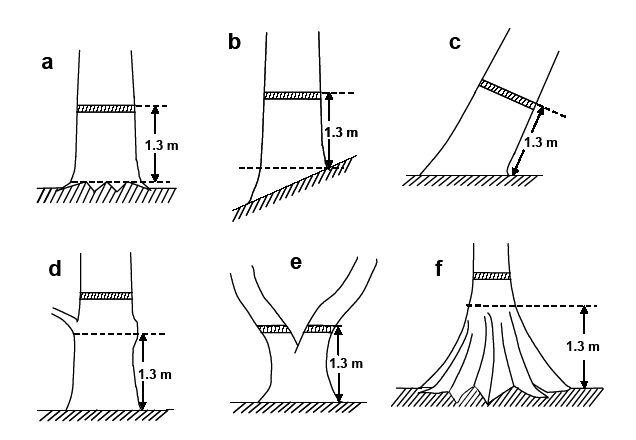
图5
胸径测量位置的确定
Fig. 5
Positions for measuring diameter at breast height (DBH) in different situations
①总是从上坡方向测定(图5b);
②对于倾斜或倒伏的个体, 从下方而不是上方进行测定(图5c);
③如树干表面附有藤蔓、绞杀植物和苔藓等, 需去除后再测定;
④如不能直接测量DBH(如分叉、粗大节、不规则肿大或萎缩), 应在合适位置测量(图5d), 测量点要标记, 以便复查;
⑤胸高以下分枝的两个或两个以上茎干, 可看作不同个体, 分别进行测量(图5e);
⑥对具板根的树木在板根上方正常处测定(图5f), 并记录测量高度; 倒伏树干上如有萌发条, 只测量距根部1.3 m以内的枝条;
⑦极为规则的树干, 应主观确定最合适的测量点, 并标记和记录测量高度。
(5) 树高测定: 树高的测定较困难。一般要求每个径级都要测定若干个体, 以使建立的树高与DBH之间的关系能够代表群落的整体情况。一般来说, 树高的测量株数应是DBH测量株数的1/3以 上。
测定树高的方法有多种。在众多的测高器中, 以日本产的伸缩式测高器最为精确。它的测量原理很简单, 实际上就是一把可收缩的尺子, 有若干节, 每节约1 m, 上刻有刻度; 内节最细, 为到达树梢部分; 外节最粗, 在外层。平时测高器收叠起来, 仅1.2 m左右 (图6)。测高器一般有10 m、12 m、15 m和20 m等规格, 因此, 20 m左右的树高均可精确测量。在郁闭和高大的林分中测量时, 一般需要两人, 一人使用测高器, 另一人在合适位置确认测高器是否到达树梢。这种伸缩式测高器的缺点是测量速度较慢, 抽出时也较费力; 另外, 测量时, 一些细小枯枝易卡在节间, 容易使测高器损坏。
图6

图6
树高测定实例照片
Fig. 6
A photo illustrating measurement of tree height in the field
在使用其他测高器如角规式测高器时, 需要注意坡度的校正。角规式测高器在国内外使用较广泛, 本文不作介绍。
3.2.5 灌木层调查
(1) 选取样方对角的两个样格(图4), 对灌木层进行详细调查。逐株(丛)记录种名、高度、株数、基径等。测量个体包括灌木种和未满足乔木层测量标准的更新幼树。
(2) 在其中一个样格内收获灌木层地上生物量、称取鲜重, 并取样带回实验室烘干称重。
(3) 在剩余的样格中, 搜寻在两个灌木样格中未出现的灌木种(包括更新幼树、苗), 记录种名。
3.2.6 草本层调查
(1) 在样方四角和中心设置5个1 m×1 m的小样方, 小样方编号方式见图4。连续监测样方须以木桩标记草本小样方的位置。
(2) 在每个草本小样方内, 记录所有草本维管植物的种名、平均高度、盖度和多度等级。
(3) 在其中两个1 m×1 m小样方内收获草本层地上生物量和地表枯落物、称取鲜重, 并取样带回实验室烘干称重。
(4) 在每个样格中, 仔细搜寻在草本小样方中未出现的草本物种, 记录种名。
3.2.7 土壤调查
在样方附近挖土壤剖面1-2个, 记录土壤剖面特征, 并以100 cm3 的土壤环刀, 按0-10 cm、10-20 cm、20-30 cm、30-50 cm、50-70 cm、70-100 cm的土壤深度分层取样, 称取鲜重并编号, 用于实验室理化性质分析。
3.3 灌丛和草地植物群落清查
全面清查研究区灌丛和草地的群落类型、物种组成、生境特征和季相等, 样方面积为10 m×10 m。具体的调查步骤和技术规范如下。
3.3.1 样方地点选择及样方设置
样方地点的选择原则参考森林群落调查。样方面积100 m2, 周围应留有10 m缓冲区, 在样方四角和中心各设置1 m× 1 m的小样方1个(图7)。
图7
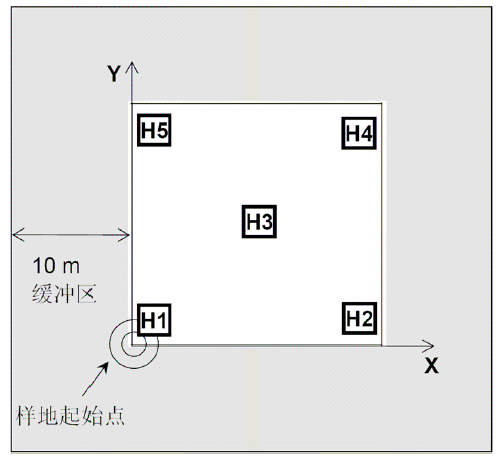
图7
灌丛(草地)样方设置方法。样方面积10 m × 10 m, 其中H1-5为详细调查小样方。样方四边应各留有10 m的缓冲区。对于灌丛, 需要调查整个样方(10 m × 10 m); 对于草地, 一般只调查5个小样方。
Fig. 7
Plot setting of shrub (grassland) communities. Five subplots (H1-5) were selected within each 10 m ×10 m plot. A buffering zone of 10 m at each side of the plot is necessary to keep the plot away from apparent human activities. For shrubs, whole plot (10 m × 10 m) should be investigated, and for grasslands, five subplots will be investigated.
3.3.2 样方环境因子调查
(1) 经纬度、海拔、坡度、坡向等测定: 同森林群落调查。
(2) 群落概况记录: 包括群落类型, 群落垂直结构, 各层次高度、盖度和优势种, 干扰和季相等。
(3) 其他样方信息的记录详见附表1。
3.3.3 样方调查
(1) 记录所有维管植物的种名、平均高度、盖度和多度等级。对灌丛, 调查整个样方(10 × 10 m); 对草地, 调查每个1 m×1 m的小样方。
(2) 在其中3个1 m×1 m小样方内收获地上生物量, 称取鲜重, 并取样带回实验室烘干称重。
(3) 在整个10 m×10 m样方内, 仔细搜寻在5个1 m×1 m小样方中未出现的物种, 记录种名。
3.3.4 土壤调查及其他
同森林群落调查。
3.4 水生植物群落清查
调查的样方面积依不同的水生植物群落而异, 一般为0.5 m×0.5 m至2 m×2 m。具体的调查步骤和技术规范如下:
3.4.1 样方设置
不同类型的水体(湖泊、河流、水库、塘堰和海洋等), 样方的设置有所不同。
(1) 湖泊水生植物群落调查: 按湖泊大小和水位, 根据等深线设置环状样带, 再根据离岸线距离设置纵向样带, 形成蜘蛛网式结构, 在每个交点处确定样方位置。
环带设置方法: 分别在沿岸带、亚沿岸带、敞水带和中央深水带设置若干条样带, 每两个样带间距离根据具体情况而定, 一般以湖底坡度比降大于0.1%确定。
纵带设置方法: 根据湖岸线形状和湖湾大小从沿岸带到中央区进行样带布设, 一般离岸距离较远则样带较少, 以免在中央区样带重叠或距离太近。
(2) 河流水生植物群落调查: 从河流源头、上游、中游、下游和河口等不同地段根据不同情况设置断面, 在每个断面上等距离设置样方。
此外, 对水库可按功能区(入水口、库区、湾汊、下泄区), 对海洋可按岸线向大洋区延伸线(红树林、潮间带、大陆架、深海区)来确定样方设置。
3.4.2 样方调查
(1) 记录样方的经纬度、海拔、水深、水温; 对于河流调查, 还需记录水流、流速、流量等因子。
(2) 在设定的样方中, 分别记录种名, 物种的物候期、高度、盖度、多度、频度及生物量, 并对各物种的不同器官(根、茎、叶)生物量进行统计。
(3) 群落高度分为自然高度和实际高度, 在挺水植物群落中两者一致; 但在沉水植物和浮叶植物群落中自然高度指水深所决定的群落高度, 而实际高度为将植物体拉直的高度。
(4) 群落生物量取样面积: 高大挺水草本植物群落取样面积为2 m×2 m; 浮叶和大型沉水植物群落取样面积为1 m×1 m; 漂浮和低矮小型沉水植物群落取样面积为0.5 m×0.5 m; 严重退化水体和植被稀疏生境的水生植物群落, 调查的面积应较大, 如10 m×10 m。
(5) 群落生物量测定: 需分别测定湿重、鲜重、风干重和烘干重。湿重为从水中取样后直接称重; 鲜重为从水中取样后停留一段时间, 待植物体表面水珠经风吹消失后称重; 风干重为从水中取样后, 将植物样品带回或原地经日光曝晒, 植物体内水分基本消失后称重; 烘干重为从水中取样后将植物样品带回实验室放烘箱中在80℃条件下烘干48 h, 植物体内水分完全消失后称重。
至于生物量取样技术, 过去多用水草夹、耙草器或刀具收割。但此法误差较大, 难以采全样方内的全部植株, 植物体不完整, 尤其根部和茎的下半段很难取到。人工下潜设置样方可以克服这些问题。设置样方时, 首先严格框定样方边界, 对较深的水体, 需要潜入水中, 将植物体扶正, 确定样方边界并将样方框好, 将漂浮样方上的外来植物体去除, 然后将样方内植株连根拔起(图8)。此法与器械收割法相比, 测定的生物量约增加10-15%。
图8

图8
深水水生植物群落的样方设置
Fig. 8
Plot setting of aquatic plant communities in deep water areas
4 群落优势种生态属性的测定
物种生态属性联系着群落物种组成和生态系统功能, 是群落对环境适应策略的反映, 是群落生态学与生理生态学结合的重要方面。近年来发现, 以功能属性为基础的“功能多样性”比“物种多样性”更能反映植物及生态系统的功能(Petchey et al., 2004; De Deyn et al., 2008), 因此测定植物的生态属性十分重要。根据功能属性的变化来研究生物多样性的功能, 以及预测植物对未来全球变化的响应是群落生态学的一个重要发展方向。
近年来, 植物属性的研究从区域到全球, 从不同类群到整个被子植物都取得了重要进展。例如, Wright等(2004)基于全球各个植被类型175个样点2,500余种植物叶属性的综合分析, 阐明了这些功能属性间普遍存在的相关规律, 发现环境对叶功能的影响非常有限, 提出了“叶经济学谱系”(leaf economics spectrum)的概念。植物叶片属性间的相互关系, 不仅有着生理生化基础, 而且与植物对环境适应的权衡(trade-off)有关(He et al., 2009)。最近的研究表明, 木材和种子的生态属性也表现出类似的规律(Moles et al., 2005; Chave et al., 2009)。
为了反映植物群落的主要功能特征和一些具有重要生态和资源价值的物种的属性, 有必要确定一批重要物种, 包括群落优势植物及指示植物、资源植物、濒危植物、特有植物等, 对其生态属性进行测定。
4.1 群落优势种生态属性及其取样规范
常见的植物属性分为叶片属性、繁殖器官属性、枝干属性、根系属性等。叶片属性包括叶片大小、比叶面积(单位重量的叶面积)、叶片干物质含量、叶寿命、叶片氮和磷含量、最大光合速率、气孔导度、蒸腾速率等。繁殖器官属性包括种子大小、传播方式等。树干属性包括树皮厚度、木材密度、心材/边材比率、树干高度等。根系属性包括根系的类型、分枝情况、根级组成、比根长(单位重量的根长度)、根系的化学组成, 以及共生真菌侵染率等(表3)。这些属性特征是植物在长期适应环境过程中为获得最大光合生产力所形成的适应策略, 是群落的重要结构和功能特征参数。
表3 植物属性的计量单位、一般数值范围、要求的最小重复数和推荐的重复数。在最小重复数和推荐重复数中, 第一个数值代表个体数, 第2个数值代表器官数。此表基于Cornelissen等(2003a)、Pregitzer等(2002)以及He等(2006b)等归纳。
Table 3
| 属性 Traits | 建议的计量单位Unit suggested | 范围 Range | 最小重复数 Minimum replications | 推荐的重复数 Suggested replications | |
|---|---|---|---|---|---|
| 叶片属性 Leaf | |||||
| 叶片大小 Leaf size | mm2 | 1-106 | 5, 2 | 10, 2 | |
| 叶片厚度 Leaf thickness | mm | 5, 2 | 10, 2 | ||
| 叶片干物质含量 Leaf dry matter content | mg/g | 50-700 | 5, 2 | 10, 2 | |
| 比叶面积 Specific leaf area | mm2/mg | 2-80 | 5, 2 | 10, 2 | |
| 叶寿命 Leaf lifespan | month | 0.5-200 | 3, 12 | 10, 12 | |
| 叶片N Leaf N concentration | mg/g | 10-60 | 5, 2 | 10, 2 | |
| 叶片P Leaf P concentration | mg/g | 0.5-5 | 5, 2 | 10, 2 | |
| 最大光合速率 Maximum photosynthetic rate | μmol·m-2·s-1 | 5, 2 | 10, 2 | ||
| 气孔导度 Leaf stomatal water conductance | mol·m-2·s-1 | 5, 2 | 10, 2 | ||
| 蒸腾速率 Leaf transpiration | mol·m-2·s-1 | 5, 2 | 10, 2 | ||
| 枝干属性 Stem | |||||
| 树皮厚度 Bark thickness | mm | 5 | 10 | ||
| 树干密度 Stem specific density | mg/mm3 | 0.4-1.2 | 5 | 10 | |
| 心材/边材比率 Heartwood to sapwood area ratio | unitless | 5 | 10 | ||
| 树干高度 Stem height | m | ? -100 | 10 | 25 | |
| 根系属性 Root | |||||
| 类型 Root type | cat. | 5 | 10 | ||
| 比根长 Specific root length | m/g | 10-500 | 5, 10 | 10, 10 | |
| 细根直径 Fine root diameter | mm | 5, 10 | 10, 10 | ||
| 组织密度 Tissue density | mg/mm3 | 5, 10 | 10, 10 | ||
| 细根N Fine root N concentration | mg/g | 5, 2 | 10, 2 | ||
| 细根P Fine root P concentration | mg/g | 5, 2 | 10, 2 | ||
| 繁殖属性 Reproduction | |||||
| 种子大小 Seed mass | mg | 10-3-107 | 3, 5 | 10, 5 | |
| 传播类型 Dispersal mode | cat. | 3 | 3 | ||
| 萌枝能力 Resprouting capacity | unitless | 0-100 | 5 | 25 | |
有关植物属性的测定, 国际上已经有比较成熟的规范, 如比较综合的测定手册(Cornelissen et al., 2003b)、叶片化学计量学的测定方法(He et al., 2006b)、根系属性的测定(Pregitzer et al., 2002)等。
表3 给出了我们推荐测定的植物属性的计量单位和取样数。由于植物叶片属性随光照、冠层位置有较大的变化, 取样时以采集冠层位置的阳生叶为最佳。
4.2 生态属性的测定
表4列出了常见生态属性的测定方法。由于生态属性的测定需要花费巨大的人力物力, 因此一定要设计出适合当地条件的测定规程。同时, 要注意季节变化的影响, 尽量在同一物候期进行测定。
表4 植物生态属性的常见测定方法
Table 4
| 属性 Traits | 测定方法 Measuring methods |
|---|---|
| 叶片属性 Leaf 叶片大小 Leaf size | 叶面积仪(如Delta-T (Cambridge, UK), Li-Cor (Lincoln, Nebraska, USA)), 或扫描仪 |
| 叶片厚度 Leaf thickness | 显微镜或游标卡尺测量 |
| 叶片干物质含量 Leaf dry matter content | 叶片装入自封袋, 加水浸湿, 保持12 h以上, 吸掉多余水分, 称鲜重; 60℃烘干至少72 h(或80℃至少48 h), 称干重, 计算 |
| 比叶面积 Specific leaf area | 面积测定: 叶面积仪(如Delta-T (Cambridge, UK), Li-Cor (Lincoln, Nebraska, USA)。干重测定: 烘箱60℃烘烤至少72 h(或80℃至少48 h), 称干重。由叶面积和干重计算比叶面积。 |
| 叶寿命 Leaf lifespan | 双子叶植物: 叶片标记法; 单子叶植物: 测定叶片特定区域的生产和死亡动态, 具体见Cornelissen 等(2003b) |
| 叶片N Leaf N concentration | 元素分析仪法(2400Ⅱ CHNS/O Elemental Analyzer, Perkin-Elmer, Boston, MA,USA)(He et al., 2006a)或凯氏定氮法 |
| 叶片P Leaf P concentration | 钼蓝比色法 (Kuo, 1996; He et al., 2008) |
| 最大光合速率 Pnmax | Li-6400便携式光合仪测定, 建议设光照=1,500 μmol/(m2·s1), [CO2] = 380 ppm |
| 气孔导度 Gs | Li-6400便携式光合仪测定 |
| 蒸腾速率 Tr | Li-6400便携式光合仪测定 |
| 枝干属性 Stem | |
| 树皮厚度 Bark thickness | 游标卡尺测量 |
| 树干密度 Stem specific density | 体积替代法: 鲜树干完全浸于盛水的量筒中约5 s, 读取增加的体积; 烘箱中60℃烘烤至少72 h (小树干)或96 h (大树干), 称干重。由体积和重量计算树干密度 |
| 心材/边材比率 Heartwood to sapwood area ratio | 钻取生长芯, 经打磨处理后, 用高精度扫描仪进行图像扫描, 据心材和边材颜色区分其界限, 用年轮分析软件测量每个生长芯的树皮厚度、边材宽度和边材年轮数。以树干横断面为标准圆形计算心材/边材比率 |
| 树干高度 Stem height | 直接测量法(如激光测距仪)和三角函数法 |
| 根系属性 Root | |
| 类型 Root type | 参照Weaver(1958)或Cannon (1949) |
| 比根长 Specific root length | 解剖镜下区分出活的、完整的根, 用目镜测微尺测量长度; 或用WinRHIZO根系分析系统, 60℃下至少烘烤72 h(或80℃至少48 h), 称重, 然后计算比根长 |
| 细根直径 Fine root diameter | 解剖镜下区分出活的、完整的根, 用目镜测微尺校正细根直径 |
| 组织密度 Tissue density | 根干重/根体积, 详细内容参见Ryser (1996) |
| 细根N Fine root N concentration | 元素分析仪法(如CHNS/O Elemental Analyzer, Perkin-Elmer, Boston, MA, USA)或微量凯氏定氮法 |
| 细根P Fine root N concentration | 钼蓝比色法 (Kuo, 1996; He et al., 2008) |
| 繁殖属性 Reproduction | |
| 种子大小 Seed mass | 野外采成熟果实, 清理(洗)出种子, 统计种子数目, 80℃下至少烘烤48 h, 直到恒重, 称重。 |
| 传播类型 Dispersal mode | 记录所有与传播有关的分类学特征, 据重要性按降序排列。具体分类特征见Cornelissen等(2003b) |
| 萌枝能力 Resprouting capacity | 根据萌枝数, 划分等级 |
5 大样地调查
大样地调查是近年来国际上兴起的一种森林植物群落学的长期研究途径, 但它本质上不属于传统意义上的群落研究, 因为在较大的连续地段(大于5 ha)上常常发育着多个群落类型。这种情况在地形较为破碎的中国尤其明显。但大样地调查对于了解景观和区域尺度的群落分布及其与环境因素的关系有着十分重要的意义。
最早的一个50 ha大样地主要由美国Hubbell设计(Hubbell & Foster, 1983), 于1980年在巴拿马Barro Colorado Island(简称BCI)的原始热带季雨林中实施。自1982年首次完成调查以来, 研究人员每5年对BCI样地进行复查。迄今, 基于BCI样地的研究, 已经发表了一系列的重要成果。这些成果不仅深化了对热带森林群落结构、动态及生物多样性的认识, 而且对近20年来植物群落学理论方面的发展, 如中性理论(Hubbell, 2001, 2006)、林窗干扰(Hubbell et al., 1999; Sheil & Burslem, 2003)、种子扩散和补充限制(Dalling et al., 2002)、种–面积关系和β多样性(Condit et al., 1996, 2002)等方面也都起到了重要的推动作用。此后, 以BCI为模板, 人们在全球热带地区先后建立了超过30个面积在16-50 ha的大型永久森林群落样地。以此为基础, 成立了热带森林科学中心(Center for Tropical Forest Science, CTFS;
经过长期探索实践, 大样地的设计、调查和科研管理已经形成了一套独特的方法体系。BCI的主要设计参与者和负责人, Smithsonian热带森林研究所的R. Condit于1998年出版了“Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots”一书, 详细介绍了大样地的设计和调查方法。这一方法体系已经成为全球大样地设计的通用技术手册。目前我国正在积极开展的森林大样地调查也以BCI样地的技术方案为蓝本(叶万辉等, 2008; 祝燕等, 2008)。
大样地调查的主要方法内容包括选址、样地调查、数据库建设与管理等方面。由于数据库建设和管理以室内工作为主, 涉及很多与外业调查不同的复杂内容, 因此本文主要介绍大样地调查中的野外工作内容(Condit, 1998)。
5.1 大样地调查程序
10 ha以上的大样地一般将整体分割成20 m×20 m的样方, 然后将每个样方分成16个5 m×5 m的小样方作为基本的测量单元(本文称其为“样元”)。大样地调查的具体内容主要包括: 地形测绘和样方设置、树木清查、物种分类、数据质量检查, 以及样地复查等。
5.2 地形测绘和样方设置
5.2.1 装备和用品供应
(1) 主要测量工具: 地质调查罗盘(或经纬仪)和50 m测量尺(或激光测距仪), 全站仪则可以替代这两项; 带有高度标志和水平调节泡、可以伸缩的测量杆尺。
(2) 20 m×20 m样方角桩: 用于永久定位。建议使用稳定、耐久的材料, 如PVC管、水泥桩等, 固定在准确位置, 标注样方号。
(3) 5 m×5 m样元的角桩: 用类似的材料进行定位标记, 有时不作永久性标记。
5.2.2 主要工作流程
(1) 确定20 m×20 m 样方网格: 仔细勘查样地地点, 选择合适的样方作为起点, 布设样方中心十字网格线; 确定、标注网格的角桩位置, 直至20 m×20 m的样方网格完全建立, 测定每两个相邻角桩之间的地形倾角; 行列坐标均从00开始, 先行后列, 设置样方标记系统。
(2) 设置5 m×5 m 样元: 将每个20 m×20 m的样方划分成16个5 m×5 m的样元, 每个样元记录采用“样方(行, 列)”表达法。
(3) 地形测量与校准: 利用两两角桩之间的测量距离和倾角, 换算成两两之间的海拔起伏, 并绘制到一定比例尺的地图上; 为方便起见, 可利用GPS测定坐标原点的海拔, 得到整体误差最小的全部样方角桩的海拔值, 并得到地形测量误差的平均值估计; 利用这些样点的坐标和海拔值, 就可以在ArcGIS中自动绘制设定比例尺的地形图。
5.3 树木清查
树木清查是大样地调查的工作主体, 包括逐一测定每个样元中的乔木、灌木, 并进行定位、挂牌标记、种类识别和胸径测量、信息记录。测量的对象为所有DBH≥10 mm的树木, 包括树蕨和棕榈植物, 但往往不考虑藤本和绞杀植物。
5.4 物种分类
大样地的物种分类非常困难, 并且往往难以同时完成。基本的目标是确认形态种, 并采集相应的标本用于鉴定。但一般要求在野外确认到属。为此, 需要在植物分类学家的参与下, 首先进行人员培训、建立工作程序和熟悉群落。
5.5 数据质量检查
由于调查队伍通常由个别专家带领绝大多数非专业人员完成, 严格的工作程序和频繁的检查必不可少, 定期抽查可减少错误和评估随机错误率。
(1) DBH数据检查: 采取交叉检验的办法, 随机选取若干个20 m×20 m的样方, 对所有DBH≥10 cm的树木和其中头两个5 m×5 m样元中所有DBH≥10 mm的所有个体进行复核。
(2) 物种分类检查: 可在样地调查完成后进行。主要采取两种方法: 一是针对样方随机取样或针对物种类群随机取样; 二是针对广布种和稀有种随机取样。
5.6 样地复查
复查的主要目的是找到以前挂牌并测定过的个体, 对新增加的DBH≥10 mm的个体进行定位、测量和挂牌, 并确认哪些个体已经死亡。
参考文献
Long-term datasets: from descriptive to predictive data using ecoinformatics
DOI:10.1658/1100-9233(2007)18[458:LDFDTP]2.0.CO;2 URL [本文引用: 1]
Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd edn
A tentative classification of root systems
DOI:10.2307/1932458 URL [本文引用: 1]
Towards a worldwide wood economics spectrum
DOI:10.1111/j.1461-0248.2009.01285.x
URL
PMID:19243406
[本文引用: 1]

Wood performs several essential functions in plants, including mechanically supporting aboveground tissue, storing water and other resources, and transporting sap. Woody tissues are likely to face physiological, structural and defensive trade-offs. How a plant optimizes among these competing functions can have major ecological implications, which have been under-appreciated by ecologists compared to the focus they have given to leaf function. To draw together our current understanding of wood function, we identify and collate data on the major wood functional traits, including the largest wood density database to date (8412 taxa), mechanical strength measures and anatomical features, as well as clade-specific features such as secondary chemistry. We then show how wood traits are related to one another, highlighting functional trade-offs, and to ecological and demographic plant features (growth form, growth rate, latitude, ecological setting). We suggest that, similar to the manifold that tree species leaf traits cluster around the 'leaf economics spectrum', a similar 'wood economics spectrum' may be defined. We then discuss the biogeography, evolution and biogeochemistry of the spectrum, and conclude by pointing out the major gaps in our current knowledge of wood functional traits.
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots, pp.1-224
Species-area and species-individual relationships for tropical trees: a comparison of three 50-ha plots
DOI:10.2307/2261477 URL [本文引用: 1]
Beta-diversity in tropical forest trees
DOI:10.1126/science.1066854
URL
PMID:11809969
[本文引用: 1]
The high alpha-diversity of tropical forests has been amply documented, but beta-diversity-how species composition changes with distance-has seldom been studied. We present quantitative estimates of beta-diversity for tropical trees by comparing species composition of plots in lowland terra firme forest in Panama, Ecuador, and Peru. We compare observations with predictions derived from a neutral model in which habitat is uniform and only dispersal and speciation influence species turnover. We find that beta-diversity is higher in Panama than in western Amazonia and that patterns in both areas are inconsistent with the neutral model. In Panama, habitat variation appears to increase species turnover relative to Amazonia, where unexpectedly low turnover over great distances suggests that population densities of some species are bounded by as yet unidentified processes. At intermediate scales in both regions, observations can be matched by theory, suggesting that dispersal limitation, with speciation, influences species turnover.
Functional traits of woody plants: correspondence of species rankings between field adults and laboratory-grown seedlings?
DOI:10.1111/jvs.2003.14.issue-3 URL [本文引用: 2]
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide
DOI:10.1071/BT02124 URL [本文引用: 3]
Role of dispersal in the recruitment limitation of neotropical pioneer species
Plant functional traits and soil carbon sequestration in contrasting biomes
The imprint of the geographical, evolutionary and ecological context on species-area relationships
DOI:10.1111/j.1461-0248.2005.00848.x
URL
PMID:16958886
[本文引用: 1]
Species-area relationships (SAR) are fundamental in the understanding of biodiversity patterns and of critical importance for predicting species extinction risk worldwide. Despite the enormous attention given to SAR in the form of many individual analyses, little attempt has been made to synthesize these studies. We conducted a quantitative meta-analysis of 794 SAR, comprising a wide span of organisms, habitats and locations. We identified factors reflecting both pattern-based and dynamic approaches to SAR and tested whether these factors leave significant imprints on the slope and strength of SAR. Our analysis revealed that SAR are significantly affected by variables characterizing the sampling scheme, the spatial scale, and the types of organisms or habitats involved. We found that steeper SAR are generated at lower latitudes and by larger organisms. SAR varied significantly between nested and independent sampling schemes and between major ecosystem types, but not generally between the terrestrial and the aquatic realm. Both the fit and the slope of the SAR were scale-dependent. We conclude that factors dynamically regulating species richness at different spatial scales strongly affect the shape of SAR. We highlight important consequences of this systematic variation in SAR for ecological theory, conservation management and extinction risk predictions.
Overestimated biomass carbon pools of the northern mid- and high latitude forests
The Ecology of Plants
Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China
DOI:10.1007/s00442-006-0425-0
URL
PMID:16639565
[本文引用: 1]
Nitrogen (N) and carbon-nitrogen (C:N) ratio are key foliar traits with great ecological importance, but their patterns across biomes have only recently been explored. We conducted a systematic census of foliar C, N and C:N ratio for 213 species, from 41 families over 199 research sites across the grassland biomes of China following the same protocol, to explore how different environmental conditions and species composition affect leaf N and C:N stoichiometry. Leaf C:N stoichiometry is stable in three distinct climatic regions in Inner Mongolia, the Tibetan Plateau, and Xinjiang Autonomous Region, despite considerable variations among co-existing species and among different vegetation types. Our results also show that life form and genus identity explain more than 70% of total variations of foliar N and C:N ratio, while mean growing season temperature and growing season precipitation explained only less than 3%. This suggests that, at the biome scale, temperature affects leaf N mainly through a change in plant species composition rather than via temperature itself. When our data were pooled with a global dataset, the previously observed positive correlation between leaf N and mean annual temperature (MAT) at very low MATs, disappeared. Thus, our data do not support the previously proposed biogeochemical hypothesis that low temperature limitations on mineralization of organic matter and N availability in soils lead to low leaf N in cold environments.
Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes
DOI:10.1007/s00442-007-0912-y
URL
PMID:18278518
[本文引用: 2]
Leaf N and P stoichiometry covaries with many aspects of plant biology, yet the drivers of this trait at biogeographic scales remain uncertain. Recently we reported the patterns of leaf C and N based on systematic census of 213 species over 199 research sites in the grassland biomes of China. With the expanded analysis of leaf P, here we report patterns of leaf P and N:P ratios, and analyze the relative contribution of climatic variables and phylogeny in structuring patterns of leaf N:P stoichiometry. Average values of leaf P and N:P ratio were 1.9 mg g(-1) and 15.3 (mass ratio), respectively, consistent with the previous observation of a higher N:P ratio in China's flora than the global averages (ca. 13.8), resulting from a lower leaf P. Climatic variables had very little direct correlation with leaf P and N:P ratios, with growing season precipitation and temperature together explaining less than 2% of the variation, while inter-site differences and within-site phylogenetic variation explained 55 and 26% of the total variation in leaf P and N:P ratios. Across all sites and species, leaf N and P were highly positively correlated at all levels. However, the within-site, within-species covariations of leaf N and P were weaker than those across sites and across species. Leaf N and P relationships are driven by both variation between sites at the landscape scale (explaining 58% of the variance) and within sites at the local scale (explaining 24%), while the climatic factors exerted limited influence (explaining less than 3%). In addition, leaf N:P ratios in two dominant genera Kobresia and Stipa had different responses to precipitation. This study suggests that geographic variation and between-species variation, rather than climatic variation, are the major determinants of grassland foliar stoichiometry at the biome level.
Taxonomic, phylogenetic and environmental trade- offs between leaf productivity and persistence
A test of the generality of leaf trait relationships on the Tibetan Plateau
The unified neutral theory of biodiversity and biogeography: a synopsis of the theory and some challenges ahead
In: Integrating Ecology and Evolution in a Spatial Context (eds Silvertown J, Amtonovics J), pp. 393-411.
Neutral theory and the evolution of ecological equivalence
DOI:10.1890/0012-9658(2006)87[1387:ntateo]2.0.co;2
URL
PMID:16869413
[本文引用: 1]
Since the publication of the unified neutral theory in 2001, there has been much discussion of the theory, pro and con. The hypothesis of ecological equivalence is the fundamental yet controversial idea behind neutral theory. Assuming trophically similar species are demographically alike (symmetric) on a per capita basis is only an approximation, but it is equivalent to asking: How many of the patterns of ecological communities are the result of species similarities, rather than of species differences? The strategy behind neutral theory is to see how far one can get with the simplification of assuming ecological equivalence before introducing more complexity. In another paper, I review the empirical evidence that led me to hypothesize ecological equivalence among many of the tree species in the species-rich tropical forest on Barro Colorado Island (BCI). In this paper, I develop a simple model for the evolution of ecological equivalence or niche convergence, using as an example evolution of the suite of life history traits characteristic of shade tolerant tropical tree species. Although the model is simple, the conclusions from it seem likely to be robust. I conclude that ecological equivalence for resource use are likely to evolve easily and often, especially in species-rich communities that are dispersal and recruitment limited. In the case of the BCI forest, tree species are strongly dispersal- and recruitment-limited, not only because of restricted seed dispersal, but also because of low recruitment success due to heavy losses of the seedling stages to predators and pathogens and other abiotic stresses such as drought. These factors and the high species richness of the community strongly reduce the potential for competitive exclusion of functionally equivalent or nearly equivalent species.
Diversity of canopy trees in a neotropical forest and implications for conservation
In: Tropical Rain Forest: Ecology and Management (eds Sutton SL, Whitmore TC, Chadwick AC), pp. 25-41.
Light-gap disturbances, recruitment limitation, and tree diversity in a Neotropical forest
Phosphorus
In: Methods of Soil Analysis. Part 3, Chemical Methods (ed. Sparks DL), pp. 869-919.
Modeling the primary productivity of the world
In: Primary Productivity of the Biosphere (eds Lieth H, Whittaker R), pp. 237-263.
The measurement of community diversity
In: Principles and Methodologies of Biodiversity Studies (生物多样性研究的原理与方法) (eds Qian YQ (钱迎倩), Ma KP (马克平)), pp. 141-165.
A brief history of seed size
DOI:10.1126/science.1104863
URL
PMID:15681384
[本文引用: 1]
Improved phylogenies and the accumulation of broad comparative data sets have opened the way for phylogenetic analyses to trace trait evolution in major groups of organisms. We arrayed seed mass data for 12,987 species on the seed plant phylogeny and show the history of seed size from the emergence of the angiosperms through to the present day. The largest single contributor to the present-day spread of seed mass was the divergence between angiosperms and gymnosperms, whereas the widest divergence was between Celastraceae and Parnassiaceae. Wide divergences in seed size were more often associated with divergences in growth form than with divergences in dispersal syndrome or latitude. Cross-species studies and evolutionary theory are consistent with this evidence that growth form and seed size evolve in a coordinated manner.
How do different measures of functional diversity perform?
Fine root architecture of nine north American trees
The importance of tissue density for growth and life span of leaves and roots: a comparison of five ecologically contrasting grasses
Vegetation-plot data and databases in Europe: an overview
Disturbing hypotheses in tropical forests
A tribute to Claude Shannon (1916-2001) and a plea for more rigorous use of species richness, species diversity and the Shannon-Wiener index
Species-area relationship in biological communities
The additive partitioning of species diversity: recent revial of an old idea
Temperature dependence, spatial scale, and tree species diversity in eastern Asia and North America
Classification of root systems of forbs of grassland and a consideration of their significance
DOI:10.2307/1931749 URL [本文引用: 1]
The worldwide leaf economics spectrum
Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Ding Hu Shan, China
Community composition and structure of Gu Tian Shan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, East China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}