|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
包头南海湿地鲤科鱼类肠道菌群与宿主生理指标的相关性
生物多样性
2025, 33 (11):
25131-.
DOI: 10.17520/biods.2025131
包头南海湿地作为重要的生态区域, 拥有丰富的鱼类资源。然而, 目前针对该湿地鱼类种群动态的长期监测数据积累不足, 相关生态学研究也较为匮乏。本研究以南海湿地6种常见鲤科鱼类鲢(Hypophthalmichthys molitrix)、鳊(Parabramis pekinensis)、鳙(Aristichthys nobilis)、翘嘴鲌(Culter alburnus)、鲫(Carassius auratus)和团头鲂(Megalobrama amblycephala)为对象, 采用高通量测序技术解析其肠道菌群结构, 结合宿主生长性能、抗氧化能力及食性生态类型特征, 探究菌群与宿主生理的关联性。结果显示: (1)共获得1,051,137条有效序列, 标准化处理后被鉴定为11,108个操作分类单元(operational taxonomic units, OTUs), 属于37门1,005属; (2)菌群组成分析显示, 6种鲤科鱼肠道中存在共同核心菌群, 其中假单胞菌门、梭杆菌门和放线菌门为共有优势菌门, 鲸杆菌属(Cetobacterium)和罗氏菌属(Rothia)为共有优势菌属; (3) LEfSe分析揭示物种特异性菌群, 如鲢肠道中蓝细菌门显著富集, 与其上层水体滤食性生态习性相关; 有20个差异菌属, 如鲢的甲基杆菌(Methylobacterium_Methylorubrum)、蓝藻类群, 鳙的邻单胞菌属(Plesiomonas)及鲫的候选竞争杆状菌(Candidatus_ Competibacter)等, 它们均与宿主的生态分布密切相关; (4)菌群结构与宿主生理指标显著相关: 肥满度(R2 = 0.731)和2,2-二苯基-1-苦基肼基(DPPH)抗氧化指标(R2 = 0.700)是主要因子, 并发现与生长、抗氧化及食性相关生物学特征的差异菌属, 如与生长性能指标显著相关的有鞘氨醇单胞菌属(Sphingomonas)、噬冷菌属(Algoriphagus)和不动杆菌属(Acinetobacter); 与抗氧化能力指标显著相关的有弧菌属(Vibrio)、气单胞菌属(Aeromonas)和邻单胞菌属; 与不同食性生态位特征显著相关的有海居菌属(Marivivens)、不动杆菌属和候选竞争杆状菌。上述结果显示, 鲤科鱼类肠道中存在共有的核心菌群, 推测这些核心菌群在短链脂肪酸生成、维生素合成及能量代谢过程中表现出功能保守性; 而物种特异性的菌群可能通过协同调控宿主营养代谢、氧化应激反应及生长相关生理过程使宿主更好地适应特定环境。研究结果为湿地鱼类资源管理和精准养殖提供了实验依据。
表3
鲤科6种鱼肠道细菌中具有潜在有益或致病功能的细菌属水平相对丰度
正文中引用本图/表的段落
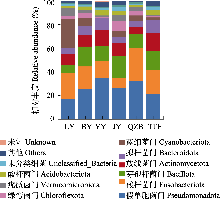
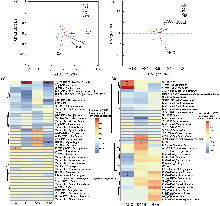
肠道微生物群落的α多样性分析显示, 鲫的Shannon指数和Simpson指数显著高于鲢(P < 0.05), 表明鲫肠道细菌群落的多样性显著高于鲢(图3c, d), 这也与鲫极强的适应性相符; 而其余组间在丰富度和多样性方面均无显著差异, 说明其他鱼类肠道细菌群落的丰富度和多样性无明显优势。基于Bray-Curtis距离的PCoA分析表明(图3e), 除LY (鲢)组外, 其余5种鱼类的样品均呈现明显的种内聚类趋势。3个主坐标轴对群落差异的解释率分别为27.41%、17.38%和9.74%。PERMANOVA检验进一步证实, 各组鱼类肠道细菌群落结构存在极显著差异(R2 = 0.529, P = 0.001)。
进一步对6种鲤科鱼类肠道细菌中的潜在有益菌和致病菌进行分析, 结果如表3所示。潜在有益菌和致病菌在不同鱼类中的分布存在差异。其中, 鲸杆菌属是丰度最高的潜在益生菌, 且在翘嘴鲌中相对丰度最高, 提示其可能在宿主营养代谢和健康维持中发挥重要作用。相比之下, 弧菌属(Vibrio)和气单胞菌属(Aeromonas)是丰度最高的潜在致病菌, 且在鳙和翘嘴鲌中相对丰度最高, 表明这些鱼类的肠道健康可能需要特别关注, 以防止潜在病原菌的过度增殖对宿主健康造成负面影响。
本文的其它图/表
|
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()