
|
|
||
|
基于类胡萝卜素着色的鸟类羽色多样性形成机制
生物多样性
2021, 29 (6):
843-854.
DOI: 10.17520/biods.2020382
人们对动物体色的研究由来已久。作为一类让生物呈现出多变色彩的重要色素, 类胡萝卜素可以在鸟类的羽毛、鸟喙和皮肤等体表组织中沉积, 产生红、橙、黄、粉、紫等颜色。类胡萝卜素不能在鸟类体内合成, 需从食物中摄取, 进而在体内完成吸收、运输、代谢和沉积等一系列过程, 才能用于羽毛着色。与类胡萝卜素着色相关的生理及遗传调控机制一直备受关注, BCO2、SCARB1和CYP2J19等影响类胡萝卜素在鸟类羽毛中着色的关键基因, 推动了对羽色遗传调控机制的深入认识。本文介绍了鸟类可利用类胡萝卜素的主要类型和基本特征, 综述了类胡萝卜素着色相关的生理过程以及调控基因研究的最新进展, 旨在增加对鸟类羽毛中类胡萝卜素着色过程和相关遗传机制的理解。 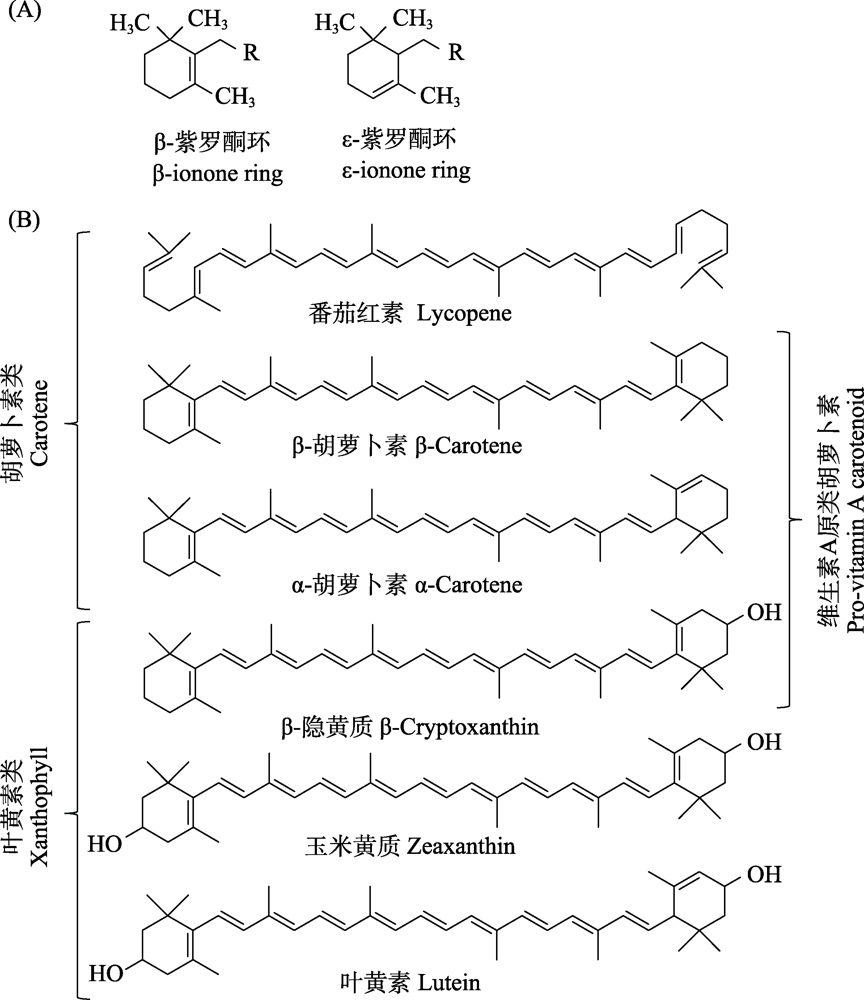 View image in article
图1
类胡萝卜素的化学结构与分类。(A)类胡萝卜素的两种端环: β-紫罗酮环与ε-紫罗酮环; (B)动物中6种具代表性的类胡萝卜素。修改自von Lintig等
正文中引用本图/表的段落
作为一类广泛存在的天然色素, 类胡萝卜素几乎完全由细菌、藻类、真菌以及高等植物合成(Olson & Owens, 1998)。动物组织中存在的类胡萝卜素属于C40四萜烯分子, 以交替的C=C双键为特征, 跨越由8个异戊二烯单元组成的碳骨架(LaFountain et al, 2015; Toews et al, 2017), 这种碳氢骨架既能以非环化的形式存在, 也可以在单侧末端或双侧末端进行环化(惠伯棣, 2005; 高广琦等, 2016)。饮食中大多数类胡萝卜素的末端都具有含1个双键的六碳环(紫罗酮环, ionone ring), 当这个双键在C5-C6位置, 则此端环为β-紫罗酮环; 当双键在C4-C5位置, 则该端环为ε-紫罗酮环(图1A) (Hill & Johnson, 2012)。
已知可用于鸟类着色的类胡萝卜素依据化学结构可分为两类: 胡萝卜素类(carotene)和叶黄素类(xanthophyll) (图1B)。第一类是纯碳氢化合物, 主要包括α-胡萝卜素(α-carotene)、β-胡萝卜素(β-carotene)和番茄红素(lycopene)等; 第二类是氧合类胡萝卜素, 主要包括叶黄素(lutein)、玉米黄质(zeaxanthin)和β-隐黄质(β-cryptoxanthin)等(Svensson & Wong,2011)。
脂蛋白被靶组织摄取是一个有选择的过程, 通常需要被靶组织(皮肤、毛囊、视网膜等)上的受体(例如SCARB1与CD36)识别, 继而以被动运输的方式被靶组织摄取, 促进类胡萝卜素向细胞内的转运(Sakudoh et al, 2013)。SCARB1可作为高密度脂蛋白受体来介导靶组织类胡萝卜素着色, 其编码基因SCARB1发生突变可能会破坏类胡萝卜素的转运过程。Toomey等(2017)发现金丝雀的白色隐性突变个体与红色或黄色的品种相比, 其组织中类胡萝卜素的含量非常低。结合遗传图谱法, 发现白色隐性等位基因是由于SCARB1的剪切供体存在点突变, 导致数量最多的剪切异构体出现异常剪切, 丧失了转运类胡萝卜素的功能, 最终导致类胡萝卜素无法在体表组织中积累着色。
本文的其它图/表
|
{kind=link}